
Abstract
Reports show that sorbitol-fermenting (SF) Escherichia coli O157 isolates are implicated in animal and human diseases and may represent new emerging pathogens. We investigated the cytotoxicity and interaction with intestinal tissues of an SF, Shiga-toxin-negative E. coli O157:NM isolate. This bovine isolate was negative for stx genes and was not cytotoxic for Vero cells. We found that this E. coli O157 isolate possesses an intimin of type β, whereas the translocated intimin receptor Tir and type III secretion system components EspA, EspB, and EspD were of type α. In contrast, Shiga-toxin-positive O157:H7 isolates usually possess variants of type γ. The isolate did not present typical O157:H7 attaching and effacing lesions in the newborn pig ileal in vitro organ culture model. However, extensive effacement and elongation of the microvilli were observed. In vitro organ culture results suggest that such an SF, Shiga-toxin-negative O157:NM isolate found in cattle may potentially cause disease, such as diarrhea without hemolytic uremic syndrome, in humans.
Introduction
Genomic differences between O157:H7 lineages were recently highlighted by ourselves and others (Dowd and Ishizaki, 2006; Lefebvre et al., 2008), and we also have reported differences in Vero cell cytotoxicity for isolates of human and bovine origin belonging to different Shiga toxin variants (Lefebvre et al., 2009). To date, however, little information is available about SF, Shiga-toxin-negative E. coli O157:H7 or non-H7 isolates, although such strains can cause human illness and may represent new emerging pathogens (Schmidt et al., 1999; Chakraborty et al., 2003). Lee and Choi (2006), as well as ourselves (Lefebvre et al., 2008), recently isolated such SF, Shiga-toxin-negative O157 strains from food production animals, which seem to also represent a reservoir for these strains. We hypothesized that such isolates may still cause host tissue damage and disease in animals and humans, and we report here the distinctive adherence of an SF O157:NM isolate to the intestinal mucosa as evaluated in the newborn pig ileal in vitro organ culture (IVOC) model.
Materials and Methods
Strain and characterization of LEE- and non-LEE-encoded genes related to T3SS
The SF, Shiga-toxin-negative O157:NM isolate (06-01-03-17) used in this study was previously identified in a rectal swab during a longitudinal study investigating the effect of growth-promoting agents and an antibiotic on the prevalence of O157:H7 in feedlot steers (Lefebvre et al., 2005). The O serotype was confirmed by slide agglutination using an O157 antiserum produced in a rabbit and by polymerase chain reaction (PCR) amplification for the O157 antigen (wbdR, wbdP, per [rfbO157], wzx, wbdO, wzy, wbdN) at the Reference Laboratory for E. coli (Faculté de Médecine Vétérinaire, Université de Montréal, St.-Hyacinthe, QC, Canada). The strain had an atypical pulse field profile, was proven negative for the fliCH7
gene (H7 antigen) by PCR, and was not motile (Lefebvre et al., 2005). A preliminary characterization of the isolate studied here previously demonstrated the presence of the following LEE-encoded T3SS variant genes: eaeA
β, tir
α, espA
α, espB
α, and espD
α (Lefebvre et al., 2008), as determined using PCR and previously published primer sequences (China et al., 1999; Garrido et al., 2006). The variant γ of these genes is usually representative of typical non-SF, Shiga-toxin-positive O157:H7 isolates (Garrido et al., 2006). In this study, PCR amplification was performed to better characterize additional LEE and non-LEE genes usually involved in the virulence of enterohemorrhagic E. coli (EHEC) or enteropathogenic E. coli (EPEC). The primers (Table 1) were designed from the sequenced genome of the O157:H7 EHEC strain EDL933 using the primer3 software (
Previously reported CGH results (Lefebvre et al., 2008).
Previously reported variant type (Lefebvre et al., 2008).
Non-LEE-encoded T3SS genes.
A smaller amplicon was generated for the atypical strain (∼600 bp) compared to that obtained for EDL933 (701 bp).
Genes previously identified as having variable sequences based on the genomic comparison of strains EDL933 and E2348/69 (Perna et al., 1998).
CGH, comparative genomic hybridization; LEE, locus of enterocytes effacement; PCR, polymerase chain reaction; NT, not tested.
Vero cell cytotoxicity
To evaluate the potential cytotoxicity of isolate 06-01-03-17, we measured the lactate dehydrogenase release by Vero cells damaged by bacterial extracts as previously described (Lefebvre et al., 2009).
IVOC model
Bacterium–intestinal cell interactions were examined using the pig ileal IVOC model previously described by Girard et al. (2005) in accordance with the Guidelines of the Canadian Council for Animal Care. Ileal segments of 15 cm from neonatal colostrum-deprived piglets were collected, and the serosa was carefully removed with forceps. Segments were longitudinally opened and mucus was gently removed. Tissues were washed in complete RPMI 1640 medium (Gibco, Burlington, ON, Canada) for 30 min on ice with constant agitation. Ileum were cut into explants of 9 mm2 and dropped with apical surface on the top on biopsy sponges placed in complete RPMI 1640 medium.
For explant infection, bacteria were grown overnight in Dulbecco's modified Eagle's medium (Gibco) at 37°C with constant agitation (180 rpm). The next day, cultures were diluted (1:50) in fresh media and incubated at 37°C until bacteria reach the exponential phase (A 600 nm of 0.7). On each explant, 50 μL of a bacterial suspension (approximately 2 × 108 CFU/mL) was dropped on the mucosal surface except for the noninfected control. Infected explants were incubated at 37°C with 5% CO2 under constant agitation for 8 h. Two hours after explant inoculation, the medium was replaced with fresh complete RPMI 1640 and subsequently changed at each hour for the rest of experimental procedure, that is, until fixation for electron microscopy.
Electron microscopy
Explants were fixed in 2.5% glutaraldehyde overnight at 4°C and then rinsed in cacodylate buffer (0.1 M cacodylate, pH 7.3) for 1.5 h with regular changes. For transmission electron microscopy (TEM), samples were postfixed in 2% osmium tetroxide at room temperature for 1 h. Tissues were then washed in water for 1.5 h with regular changes and dehydrated in successive ethanol baths.
For TEM, samples were embedded in epon-araldite resin and 80-nm sections were mounted on nickel grids. Staining was carried out with uranyl acetate and lead citrate. The samples were observed with a Philips 201 transmission electron microscope at 80 kV (Philips Electronics, Eindhover, The Netherlands).
For scanning electron microscopy (SEM), tissues were dehydrated with ethanol, dried to critical point, mounted on aluminum stub, sputter coated (10 nm) with gold, and observed with an S-3000N scanning electron microscope (Hitachi, Rexdale, ON, Canada) at 5 kV.
Results and Discussion
As expected for Shiga-toxin-negative isolates, strain 06-01-03-17 and the nonpathogenic E. coli strain K12 provoked very little lactate dehydrogenase release from Vero cells. The atypical strain was 15 and 50 times less cytotoxic than Shiga-toxin-producing O157:H7 strains and a β-hemolytic HlyA-positive E. coli, respectively (data not shown). Besides, investigation of the isolate T3SS and related genes (Table 1) suggested that genes espJ, nleA, espH, sepZ/espZ, map, and cesF were absent from the Shiga-toxin-negative O157:NM strain, whereas at least 23 other T3SS and related genes were present (eaeA, espABDFG, tir, escD [pas], escF, yhiEF, cesDT, sepQ, sepABC [escVNC], escJRSTU, and ler).
The characteristics of the studied O157:NM isolate suggest the presence of an atypical variant T3SS that may possibly alter its pattern of adherence to enterocytes as compared to that normally observed in typical A/E pathogens (EHEC and EPEC). We thus used an IVOC model to study the adherence and T3SS function of this atypical isolate. It has previously been demonstrated that porcine and human EPEC, as well as human E. coli O157:H7, isolates adhere to the epithelial cells of the porcine ileum in this model (Girard et al., 2005). Accordingly, we used a well-characterized porcine EPEC strain (ECL1001) as the A/E reference strain for this model and a well-characterized EHEC (O157:H7) strain of human origin (EDL933) as another relevant comparative strain. Further, ECL1001 possesses the same eaeA variant (β) as that of the atypical O157:NM isolate. Using electron microscopy, typical attachment and effacement lesions were observed for isolate ECL1001 (Fig. 1A, B). TEM showed intimate-adherent bacteria with microvillus effacement beneath the adherence site. Bacteria were positioned in palisades and pedestals were observed (Fig. 1A, white arrow) under adherent bacteria with some visible actin condensation (Fig. 1B, small black arrow pointing at the dark region). On SEM, we noted brush-border disorganization and an intimate contact between bacteria and the apical surface of enterocytes (Fig. 1C). Tight attachment to enterocytes and microvillus effacement were also observed for the human O157:H7 strain EDL933 (Fig. 1D, E). In contrast, A/E lesions were not observed for the atypical O157:NM bovine isolate on porcine ileal explants. Intimate attachment to epithelial cells was not frequently observed on TEM for the atypical isolate. Adherent bacteria were, nevertheless, observed with bacteria often separated from the epithelial cell surface by a gap (Fig. 1F). Remarkably, bacterial footprints accompanied by impressive microvillus effacement were frequently observed with no evidence of pedestal formation or actin polymerization. Marked brush-border reorganization and microvillus elongation were also observed on TEM and SEM (Fig. 1F, G, long black arrows).
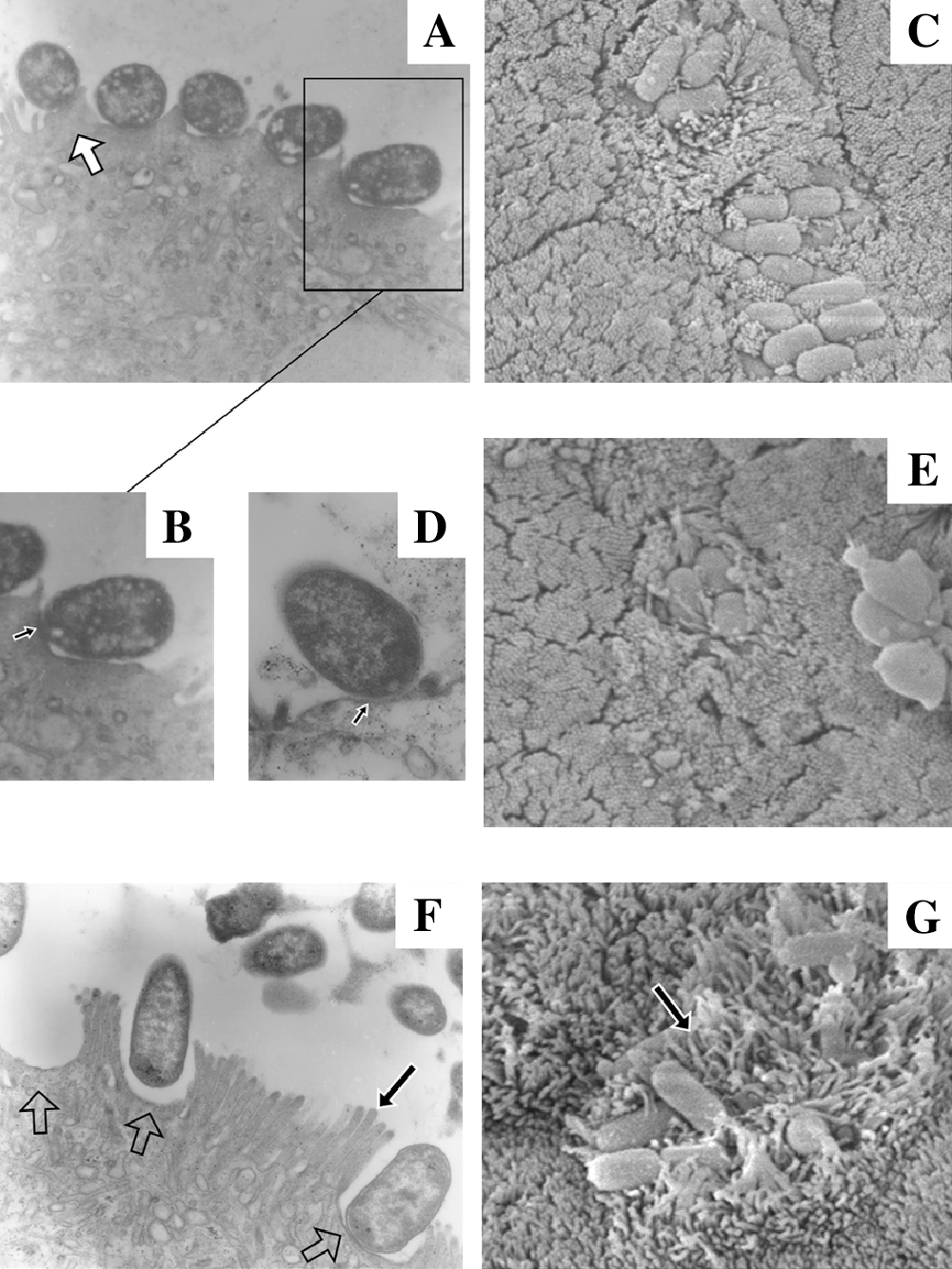
Electron microscopy of porcine ileal segments inoculated with the porcine EPEC isolate ECL1001 (
It is possible that a weaker association between the heterologous intimin (β) and Tir (α) of the atypical O157:NM isolate may explain the lack of intimate adherence between the enterocyte and the microorganism as seen here in our IVOC experiments. The absence of pedestal formation and typical A/E lesions may possibly be due to the presence of such LEE gene variants or alternatively to the absence of some T3SS effectors as suggested by our PCR analyses. Experimental validation for these hypotheses remains to be carried out. Nevertheless, the particular adherence pattern and the elongated microvilli we observed for the SF O157:NM isolate reflects its ability to adhere to the intestinal mucosa. A recent study on SF, STEC O157:NM isolates showed an increased curli-fimbriae-associated adherence of such strains to Caco-2 cells compared to non-SF O157:H7 strains. The authors suggested that this property may account for the higher incidence of disease progression to hemolytic uremic syndrome that was observed for such an emerging pathogen in Europe (Rosser et al., 2008).
To our knowledge, this is the first detailed characterization of an SF, Shiga-toxin-negative O157:NM isolate naturally occurring in the bovine gut, and which possessed a truncated LEE and an atypical pathotype (eaeA β tir α espA α espB α espD α). This isolate was positive by DNA hybridization for the curli and type I fimbriae and was also described as hypermutable (Lefebvre et al., 2008). Whether these characteristics are responsible for the particular pattern of adherence, the acquisition of the atypical T3SS variant genes, or the emergence of new pathogens, in general, remains to be investigated.
Footnotes
Acknowledgments
This work was supported by Grants 89758-01 and 2294 from the Natural Sciences and Engineering Research Council of Canada to F.M. and J.M.F., respectively, and from Agriculture and Agri-Food Canada to M.S.D. We thank Clarisse Desautels, Lisette Beaudoin, Brigitte Lehoux, and Jade-Pascale Prévost Lemyre and Heidi Rempel for technical assistance; Diane Montpetit for SEM; and Céline Lussier Lusignan for TEM.
Disclosure Statement
No competing financial interests exist.