
Abstract
The aim of this study was to evaluate a fast and simple bead-based method using paramagnetic beads covered with recombinant Listeria phage endolysin–derived cell-wall-binding domain proteins specific for Listeria spp. for separation of the foodborne pathogen Listeria monocytogenes from artificially contaminated raw milk. The method was combined with subsequent detection and quantification by the traditional plate-count technique and real-time polymerase chain reaction (PCR). To account for differences in cell properties, recovery rates and the detection limit were determined using five different L. monocytogenes strains for preparation of a 10-fold dilution series in raw milk, spanning an 8-log scale. Two independent test series were performed for each strain, yielding mean recovery rates of 46.6% to 122.8% for detection with the plate-count method, and 64.7% to 95.1% for detection by real-time PCR. A high correlation was found between the number of L. monocytogenes added to the samples and the number of colony forming units recovered by plate count (0.980), as well as the number of bacterial cell equivalents obtained by real-time quantitative PCR (0.987). The detection limit of the combined cell-wall-binding domain proteins/real-time PCR approach ranged from 102 to 103 colony forming units per milliliter, which is close to the theoretical detection limit of the method.
Introduction
L
Traditional microbiological methods for detection of L. monocytogenes in food are time consuming, including selective enrichment and plating on agar plates, followed by tests for species identification (Anonymous, 1996, 1998a). Quantification methods include a short period of preincubation followed by plating on selective agar plates (Anonymous, 1998b, 1998c). Faster detection methods targeting nucleic acids such as polymerase chain reaction (PCR) or real-time PCR were developed (Nguyen et al., 2004; Rossmanith et al., 2006; Mafu et al., 2009; Omiccioli et al., 2009). The majority of these methods are highly susceptible to interfering substances in the environment or in food samples (Hoorfar et al., 2004). A further important aspect is the concentration of target microorganisms in the sample. To address these issues, different ways for separation and concentration of bacterial targets from food have been used. Apart from unspecific methods such as filtration or centrifugation, specific methods like immunomagnetic separation using antibody-covered beads have been employed (Besse et al., 2004; Stevens and Jaykus, 2004). Several publications refer to immunomagnetic separation of L. monocytogenes from food after a preenrichment step, but only a few address the direct separation of Listeria cells from food, such as milk or cheese homogenate (Fluit et al., 1993; Nogva et al., 2000; Uyttendaele et al., 2000; Hsih and Tsen, 2001; Kaclíková et al., 2001; Gray and Bhunia, 2005; Yang et al., 2007).
Recently, paramagnetic beads coated with recombinant Listeria phage endolysin–derived cell-wall-binding domain proteins (CBD beads) specific for Listeria spp. were successfully used for the detection of L. monocytogenes in food, in combination with an enrichment step (Kretzer et al., 2007). Compared to antibodies, CBD beads have similar equilibrium constants, but offer the advantage of in vitro production thus not relying on the use of laboratory animals (Loessner et al., 2002). Bead-based separation protocols are fast, are easy to perform, and have the inherent option of automation (Prentice et al., 2006). As a prerequisite for further developments, the suitability of these beads for separation of L. monocytogenes directly from raw milk rather than from the enrichment broth was thoroughly assessed in the present study.
Combining bead-based separation of L. monocytogenes from raw milk with culture-based and real-time PCR–based quantification, recovery rates (RRs) at different target concentrations, and the detection limit were determined by analyzing dilution series of five different L. monocytogenes strains in raw milk in two independent experimental series.
Materials and Methods
Bacterial isolates and culture conditions
The strains used for inoculation of raw milk were the following: L. monocytogenes strains SLCC 2755 (serovar 1/2b, internal number 2934), SLCC 2482 (serovar 7, internal number 2935), NCTC 5105 (serovar 3a, internal number 2938), NCTC 10527 (serovar 4b, internal number 2942), and NCTC 7973 (serovar 1/2a, internal number 2945). Strains were activated overnight from −80°C stock cultures in sterile brain heart infusion broth supplemented with 0.6% yeast extract (Merck) at 37°C. A volume of 100 μL overnight culture was transferred to 8 mL brain heart infusion broth supplemented with 0.6% yeast extract and subcultured at 37°C for 18 h. A 10-fold dilution series was prepared in Ringer's solution and plated in duplicates onto tryptic soy agar plates supplemented with 0.6% yeast extract (TSA-Y; Merck). The TSA-Y plates were incubated at 37°C for 24 h. The colonies were enumerated and the final concentration was expressed as CFU/mL. All strains were obtained from the collection of bacteria at the Institute of Milk Hygiene, Milk Technology and Food Science, Department of Veterinary Public Health and Food Science, University of Veterinary Medicine, Vienna, Austria.
Raw milk for artificial contamination
Raw milk was obtained from a local dairy farm of the University of Veterinary Medicine, Vienna, Austria. It was not heat treated, thus containing the natural raw milk microbiota. The same batch of raw milk was used for all experiments. A 100-μL volume of the L. monocytogenes subculture was added to 9.9 mL raw milk. Raw milk samples containing 106–10−1 CFU of L. monocytogenes per mL were prepared by 10-fold dilutions in raw milk. Raw milk not inoculated with L. monocytogenes served as negative control and was subjected to the same experimental procedure as described below and tested negative.
CBD beads for recovery of L. monocytogenes from raw milk
CBD dynabeads M-270 epoxy were prepared at the Institute of Food Science and Nutrition, Swiss Federal Institute of Technology, Zurich, Switzerland. In accordance with recommendation of Kretzer et al. (2007), 10-μL beads (2 × 107 beads) were mixed with 100 μL of artificially contaminated raw milk containing 106 to 10−1 CFU of L. monocytogenes/mL, and adjusted to a final volume of 200 μL with 1 × phosphate-buffered saline containing 0.1% Tween 20 (PBST; pH 8). The samples were incubated for 2 h in an overhead rotator at 10 rpm (Dynal Sample Mixer, Model MXIC 1; Invitrogen) at room temperature, followed by separation with a magnetic separator (Magnetic Particle Concentrator for Eppendorf microtubes; Dynal). After discarding the supernatant, the beads were washed twice with 1 mL PBST.
Detection and quantification of L. monocytogenes recovered from raw milk
Bacteria recovered on the beads were (1) plated onto Palcam agar plates (Biokar Diagnostics) and incubated for 24–48 h at 37°C and (2) subjected to DNA isolation and real-time PCR analysis.
For spread plating, the beads were suspended in a 1-mL volume of PBST. A 10-fold dilution series was prepared in Ringer's solution and plated onto Palcam agar plates. The plates were incubated at 37°C for 48 h. The colonies were enumerated and the result was expressed as CFU/mL.
For DNA isolation the beads were directly subjected to the NucleoSpin® tissue kit (Clontech − Takara Bio Europe) and the support protocol for Gram-positive bacteria. The technology of this kit is based on DNA binding to silica in the presence of chaotropic salt.
Real-time PCR was performed in an Mx3000p real-time PCR thermocycler (Stratagene). Primers were used to amplify a 274-bp fragment of the prfA gene specific for L. monocytogenes (Rossmanith et al., 2006). Forward primer (LIP1: 5′-GATACAGAAACATCGGTTGGC-3′) and reverse primer (LIP2: 5′-GTGTAATCTTGATGCCATCAGG-3′) were purchased from MWG Biotech, and the TaqMan™ probe (LIP probe 2: 5′-FAM-CAGGATTAAAAGTTGACCGCA-MGB-3′) was from Applied Biosystems.
The 25-μL PCR contained 20 mM Tris-HCl, 50 mM KCl, 3.5 mM MgCl2, 500 nM of each primer, 250 nM probe, 200 μM (each) of dATP, dTTP, dGTP, and dCTP, 1.5U Platinum® Taq DNA Polymerase (Invitrogen), and 5 μL bacterial DNA. Amplification was performed after denaturation at 94°C for 2 min, with 45 cycles each at 94°C for 15 s and 64°C for 1 min.
For the quantification standard, DNA was isolated from 1 mL of an overnight culture of L. monocytogenes using the NucleoSpin tissue kit as described above. The DNA concentration was measured using the minor-groove-binding DNA stain bisbenzimide trihydrochloride (Hoechst 33258; Invitrogen) and a Hoefer DyNA Qant200 fluorometer (Pharmacia Biotech). The copy number was calculated on the basis of the molecular mass of a single copy of the genome of L. monocytogenes, as described previously (Rossmanith et al., 2006). Real-time PCR–derived target numbers were expressed as bacterial cell equivalents (BCE).
Statistical analysis
Statistical analysis was carried out using the software package SPSS 13.0 (SPSS). Data were checked for normal distribution using the Kolmogorov–Smirnov test of goodness of fit. Differences between normally distributed data sets were tested with the paired-samples t-test. RRs for bacterial cells were calculated using the equation RR (%) = x/(y/100), with x being the estimated number of CFU or BCE recovered from the beads, and y being the number of CFU inoculated into a given sample, which was determined on TSA-Y plates as described above.
Results
Direct quantification of L. monocytogenes in raw milk
The mean RRs determined on Palcam agar plates for strain 2934, 2935, 2938, 2942, and 2945 were 120.5%, 46.6%, 122.8%, 104.2%, and 88.1%, respectively. The mean RRs determined by real-time PCR for strain 2934, 2935, 2938, 2942, and 2945 were 93.0%, 69.6%, 64.7%, 95.1%, and 93.7%, respectively. Thus, a mean RR of 97.77% (RSD, 60.37) was achieved on Palcam agar plates, and 70.1% (RSD, 48.4) for real-time PCR detection. When comparing RRs from the different contamination levels, it was found that more bacterial cells were recovered at higher dilution levels (Fig. 1).
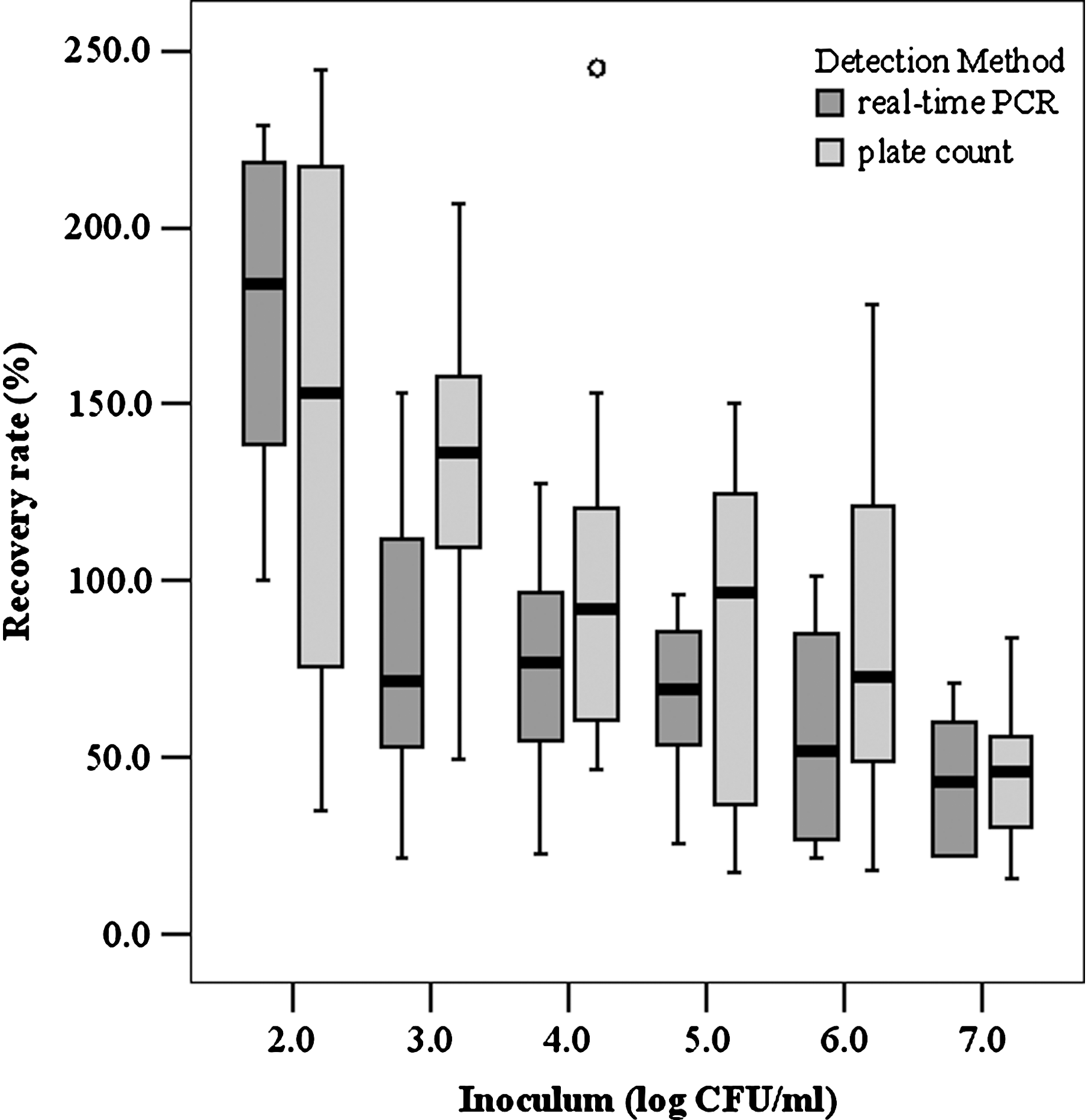
Influence of cell concentration on the recovery rate of Listeria monocytogenes from raw milk using either plate-count method or real-time polymerase chain reaction (PCR) for detection of bead-recovered cells.
The correlation of the CFU recovered on Palcam agar and the inoculum for strain 2934, 2935, 2938, 2942, and 2945 was 0.997, 0.996, 0.994, 0.970, and 0.964, respectively, with a mean correlation of 0.980 (Fig. 2). For BCE determined with real-time PCR, the correlation of these data with the inoculum for strain 2934, 2935, 2938, 2942, and 2945 was 0.989, 0.977, 0.994, 0.997, and 0.985, respectively; the mean correlation was 0.987 (Fig. 3). However, when comparing the paired data, a significant difference between (1) the number of CFU and the inoculum (p = 0.025) and (2) the number of BCE recovered on the beads and the inoculum was observed (p < 0.0005).

Correlation of inoculum and cell-wall-binding domain proteins (CBD) bead-based viable counts of colony forming units (CFU) from raw milk on Palcam agar plates.
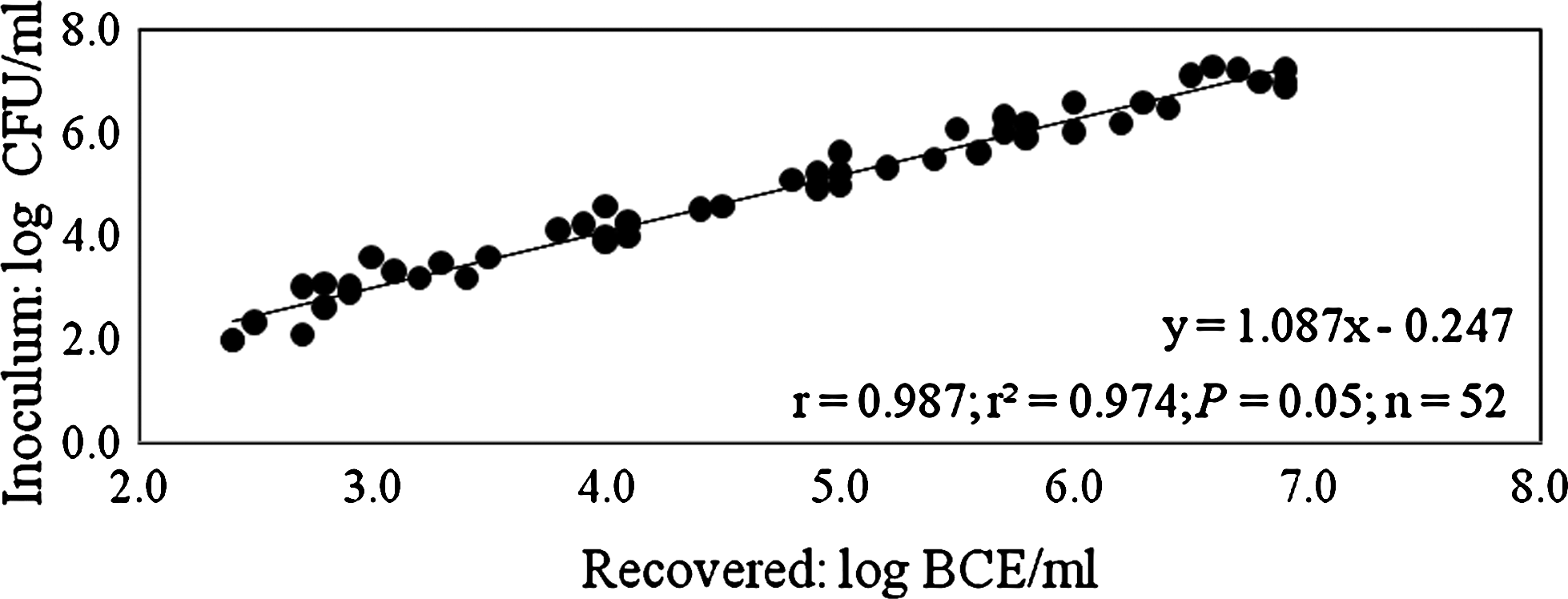
Correlation of inoculum and CBD bead-based determination of bacterial cell equivalents (BCE) from raw milk detected with real-time PCR.
The detection limit of the combined beads/real-time PCR method ranged from 102 to 103 CFU/mL raw milk (Table 1).
CFU, colony forming units.
Discussion
In most cases, immunomagnetic separation of Listeria spp. from food followed by culture-, PCR-, or cytotoxicity-based detection was combined with a selective preenrichment step (Fluit et al., 1993; Hsih and Tsen, 2001; Gray and Bhunia, 2005). Kaclíková et al. (2001) used centrifugation for the separation of nonmagnetic bead–bacterial complexes from cheese after enrichment in half-Fraser broth. The CBD beads tested in the present study have also been used successfully for culture-based detection of L. monocytogenes in different food samples after enrichment and paramagnetic separation (Kretzer et al., 2007). Few publications report isolation of L. monocytogenes with immunomagnetic beads directly from milk (Nogva et al., 2000; Yang et al. 2007). None of these included an equivalent elaborate experimental setup to collect sound performance data as we did in the present study: five different L. monocytogenes strains, 10-fold dilution series covering 8-log scales, and two independent test series for each strain.
RRs achieved with the CBD beads in the present study were 97.8% for detection on Palcam agar plates and 70.1% for detection by real-time PCR (Fig. 1). The higher RRs registered for plate-count data compared to real-time PCR data might have been due to DNA loss during isolation. The RRs compare favorably with those obtained by Yang et al. (2007) using immunomagnetic nanoparticles for separation of L. monocytogenes from milk (agar plates: 1.9% to 6.6% recovery; real-time PCR: 8% to 9% recovery for samples containing 107 to 104 CFU/mL and 27.4% and 133.4% for samples containing 103 and 102 CFU/mL, respectively). RRs were lower when commercial anti-Listeria beads were used: 0.1% to 1.8% for culture-based data and 0.1% to 0.4% for real-time PCR–based data (Yang et al., 2007). More data concerning RRs of immunomagnetic separation have been derived from pure cultures. RRs of 7% to 23% for polyclonal antibodies (Jung et al., 2003) and 1.4% to 19% for artificially created single-chain antibody fragments (Paoli et al., 2007) have been reported using plate count for enumeration.
RRs varied at different cell concentrations, yielding higher values at lower contamination levels (Fig. 1). Others reported a similar trend when using immunomagnetic nanoparticles combined with plate-count method or real-time PCR (Yang et al., 2007). The higher RRs for lower inoculum levels might have been due to a higher (and therefore better) bead-to-cell ratio. Thus, increasing the bead count might further enhance RRs at higher inoculums levels.
RRs of more than 100% were observed at the lowest contamination levels for plate count as wells as real-time PCR data (Fig. 1). These results might be due to a bias in culture-based estimation of the inoculum level, as one colony might emerge from more than one cell (Hein et al., 2005). On the other hand, cells might have been replicating during the incubation step on the rotator at room temperature. Concerning real-time PCR data, the presence of more than one genome in rapidly replicating cells could be contributing to this phenomenon (Lewin, 1997).
Given that five different strains were tested independently two times, the correlation between the number of L. monocytogenes added and the number of CFU (0.980; Fig. 2) and BCE (0.987; Fig. 3) recovered from beads was remarkable. However, a significant difference was registered between inoculums and bead-recovered CFU and BCE, as recovery was less than 100% in most cases.
The detection limit achieved experimentally ranged from 102 to 103 CFU/mL raw milk (Table 1). Samples containing less than 102 CFU/mL raw milk did not yield a positive result. The theoretical detection limit of the combined beads/real-time PCR method is at least 200 targets/mL milk. This was deduced (1) from the volume of analyzed milk (100 μL), (2) the volume of DNA solution recovered from the analyzed milk (100 μL), and (3) the volume of DNA solution analyzed in the real-time PCR assay (5 μL). As demonstrated earlier, it was assumed that the real-time PCR method is able to detect a single target if it is present in the reaction (Rossmanith et al., 2006). If one target is found in the analyzed 5-μL sample volume, it would mean that 200 targets would be present per milliliter of milk. However, as low target numbers are not distributed normally but according to Poisson's distribution, more than 200 targets/mL would have to be present in the milk to ensure that each sampled 5-μL aliquot would actually contain a target, thus yielding a positive signal (Berdal and Holst-Jensen, 2001; Hein et al., 2005). Taking into account these aspects and the fact that the RR was lower than 100%, the achieved detection limit of 102–103 CFU/mL milk lies in the anticipated range.
With respect to data reported by other authors for antibody-based separation, the detection limit achieved (102 to 103 CFU/mL raw milk) is comparable or superior. Depending on the type of beads used, detection limits ranging from 2 × 102 to 2 × 105 CFU/mL were reported for separation of L. monocytogenes from milk with immunomagnetic beads, followed by real-time PCR quantification (Nogva et al., 2000). Yang et al. (2007) achieved positive results with immunomagnetic nanoparticles for separation and real-time PCR detection for milk samples containing at least 102 CFU of L. monocytogenes/0.5 mL and 105–106 CFU of L. monocytogenes/0.5 mL when using commercial anti-Listeria beads.
The detection limit might be further lowered by subjecting larger volumes of milk to the bead-based separation protocol, concentrating the isolated DNA to get more DNA into the PCR, or combining the two approaches. This would enable detection and quantification of less than 102 CFU of L. monocytogenes/mL milk, which is an important issue for legal purposes when considering the cutoff value of 102 CFU/g for some food types (Anonymous, 2005). Since the method performed well with respect to quantification also, further experiments involving repeatability and reproducibility testing could be performed to explore the quantification limit.
Experiments with heat-treated cultures suggest that the CBD beads are unable to selectively separate viable Listeria cells (data not shown). Similar problems also arise when using antibodies (Zhou and Jiao, 2005). Recently, ethidium and propidium iodide were applied to block the amplification of DNA from dead cells and free DNA (Nocker and Camper, 2006). Preliminary experiments with propidium iodide treatment of milk before bead-based separation have been promising and should be investigated further.
Conclusions
The CBD bead separation method is a fast and simple tool for determining concentrations of L. monocytogenes in raw milk, and can be combined with culture or real-time PCR analysis. The CBD beads used in the present study yielded excellent RRs over a wide range of concentrations with either analysis method. The beads might be combined with an automated recirculating immunomagnetic capture system, similar to the AOAC-validated method currently in use for pooling and concentrating Escherichia coli O157:H7, Salmonella spp., or Listeria spp. (Prentice et al., 2006).
Footnotes
Acknowledgment
This work was supported by the European Union–funded integrated project BIOTRACER (contract 036272) under the 6th RTD.
Disclosure Statement
No competing financial interests exist.