
Abstract
Noroviruses and hepatitis A virus (HAV) are common causes of foodborne disease. They are usually shed in feces and have been found in sewage water, biosolids, and animal manures. With the wide application of manure and biosolids on agricultural lands, there is an increasing interest in investigating virus survival in manure and biosolids. In this study, Murine norovirus-1 (MNV) and HAV were inoculated into different types of animal manure and three types of differently treated biosolids at 20°C and 4°C for up to 60 days. Both HAV and MNV viral genomes degraded immediately in high pH biosolids type 2 and 3 at time zero. For other types of manure and biosolids, HAV RNA was significantly reduced in biosolids type 1 and in liquid dairy manure (DM) after 60 days stored at 20°C, but was stable in all types of manure and biosolids type 1 at 4°C. MNV RNA was unstable in pelletized poultry litter and biosolids type 1 at 20°C, and less stable in liquid DM at both temperatures. For MNV infectivity, there was no significant difference among pelletized poultry litter, alum-treated poultry litter, raw poultry litter, and swine manure at either 20°C or 4°C after 60 days of storage. However, HAV stored in swine manure and raw poultry litter had significantly higher infectivity levels than HAV stored in alum-treated poultry litter at both 20°C and 4°C. Overall, both viruses were inactivated rapidly in alkaline pH biosolids and unstable in liquid DM, but alum added in poultry litter had different effects on the two viruses: alum inactivated some HAV at both temperatures but had no effect on MNV.
Introduction
N
Meanwhile, application of manure and biosolids on agricultural land is still the most desirable way to recycle the waste and improve the soil. Although there is an increasing concern regarding the consumption of fecally contaminated foods and transmission of human enteric viruses (Sair et al., 2002), little is known regarding the survival of viruses in complex matrices such as manure/biosolids and how they are transported to plants. Previous studies have reported the use of reverse transcriptase–quantitative real-time polymerase chain reaction (RT-qPCR) in detection of NoV in human stool (Richards et al., 2004) and waste water (da Silva et al., 2007) samples, and the use of immunomagnetic capture RT-PCR to detect HAV in sewage water (Jothikumar et al., 1998). However, a positive signal in these tests indicates the presence of an intact segment of viral genomic RNA, but does not provide information on virus infectivity (Koopmans and Duizer, 2004). Information on not only the quantity of viral genomes but also the infectious viruses is urgently needed for the assessment of the public health risks posed by NoV and HAV infections. The objective of our study was to evaluate the stability of not only viral genomic copies but also infectivity of murine norovirus-1 (MNV) (a widely used surrogate for human NoVs) and HAV in different types of manure and biosolids.
Materials and Methods
Viruses
MNV-1.CW1 strain was kindly provided by Dr. Herbert Virgin IV (Washington University School of Medicine, St. Louis, MO) and was propagated on confluent monolayers of RAW 264.7 cells cultured in Dulbecco's modified Eagle's medium (Gibco-Invitrogen), supplemented with 10% fetal bovine serum, 1% glutamax, 1% penicillin/streptomycin, 1% HEPES buffer, and 1% glutamine (Wobus et al., 2004). HAV, cytopathic strain HM-175 (ATCC VR-1402), was propagated in fetal rhesus monkey kidney (FRhK-4) cells in Dulbecco's modified Eagle's medium, supplemented with 1% penicillin/streptomycin, 1% HEPES buffer, and 2% or 10% fetal bovine serum, respectively, for maintenance or growth. Viruses in infected cell lysates were purified by one freeze–thaw cycle, followed by centrifugation at 2500 g for 15 min. The upper aqueous layer was recovered and stored at −80°C for further analysis. Virus titer was determined by RT-qPCR of 10-fold dilutions of virus stock to an endpoint, and the highest dilution producing a positive RT-qPCR result was defined as 1 RT-qPCR unit (RT-qPCRU). The MNV stock was ∼1 × 108 RT-qPCRU/mL, and the HAV was ∼1 × 108 RT-qPCRU/mL.
The plaque assay for MNV was conducted as described by Wobus et al. (2004) using confluent monolayers of RAW cells grown in 12-well plates. HAV infectious titer was evaluated by tissue culture infectious dose 50 (TCID50) using FRhK-4 cells grown in 96-well plates. The infectious titer of virus stock was ∼2.5 × 106 plaque forming unit (PFU)/mL for MNV and was 4 × 107 TCID50/mL for HAV.
Manure/biosolids
Three types of wastewater biosolids were kindly provided by local waste water treatment plants (WWTP). Biosolids type 1 was obtained from Back River WWTP (Baltimore, MD) and FeCl3 was added for phosphate control (reducing soluble phosphate by adsorption of it to the surface of ferric hydroxide). Biosolids type 2 was from Seneca WWTP (Laurel, MD), and was lime and alum treated. Heat- and lime-stabilized biosolids type 3 was provided by Little Patuxent WWTP (Savage, MD). The major chemical components were measured in a previously study (Shober and Sims, 2009) or provided by the WWTP (Seneca) (Table 1). Pelletized poultry litter (PL) was kindly provided by Perdue AgriRecycle (Sussex County, DE) and raw poultry litter (RPL), alum (aluminum sulfate)-treated poultry litter (AL), and liquid dairy manure (DM) were obtained from the College of Agriculture and Natural Resources farm of University of Delaware. Swine manure (SM) was obtained from a local farm in Kent County, DE. All of the manure and biosolids were autoclaved and stored at 4°C before use.
Virus survival study
One milliliter of HAV (∼1 × 107 TCID50/mL) or MNV (∼2.5 × 105 PFU/mL) was added to 2 g solid manure or biosolids, dried for ∼15 min, stored at 20°C or 4°C, and analyzed every 10 days. For liquid DM, MNV was inoculated into 1 mL liquid DM to a final concentration of 1.25 × 105 PFU/mL and HAV was added to DM to 5 × 106 TCID50/mL, and stored at 20°C or 4°C. All samples were set in duplicate for RT-qPCR analysis and culture assay. Viral RNA was directly extracted from manure solution using QIAshredder™ homogenizer (Qiagen) and QIAamp Viral RNA Mini Kit (Qiagen) and applied to two step RT- and qPCR as described below. The concentration of viral RNA in each manure sample was then obtained from standard curves, and MNV or HAV RNA sample extracted from virus spiked manure/biosolids used for standard curve generation was used as internal control for all RT-qPCR analysis. The concentration of infectious MNV and HAV was determined using plaque assay or TCID50 method, respectively.
Viral RNA extraction and standard curve generation
Ten milliliters of Na2HPO4 (0.15 M, pH 9.5) was added into each sample of the manure and biosolids samples, and vortexed for 30 min to obtain a homogeneous suspension (10−1). Then, 200 μL of suspension was transferred into the QIAshredder homogenizer and centrifuged at 16,000 g for 2 min, and the elution was used for RNA extraction. The RNA was extracted following the manufacturer's instructions using QIAamp Viral RNA Mini Kit. Briefly, 70 μL sample was mixed with 280 μL AVL buffer (guanidine thiocyanate), 2.8 μL RNA carrier, and 1 μL RNase inhibitor and incubated at room temperature for 10 min. The RNA was precipitated by the addition of 100% ethanol and further washed with AW1 (guanidine hydrochloride) and AW2 (56% ethanol) buffer with centrifugation. The RNA was eluted with AVE (RNase free water, 0.04% sodium azide) buffer and stored at −80°C.
The standard curve was generated as described by Wei et al. (2009). Briefly, 1 × 108 RT-qPCRU/mL MNV or 1 × 108 RT-qPCRU/mL HAV was added into 2 g manure/biosolids, dried for 15 min in a biosaftey cabinet, and vortexed with 10 mL 0.15 M Na2HPO4 (pH 9.5) for 30 min (10−1 suspension). The 10−1 suspension was then homogenized using the QIAshredder homogenizer and serially diluted to 10−5 with Hanks' balanced salt solution, and viral RNA was extracted from all dilutions. For liquid DM, 0.5 mL 1 × 108 RT-qPCRU/mL MNV or 1 × 108 RT-qPCRU/mL HAV was added to 0.5 mL sample and analyzed as described above. A two-step RT-qPCR assay using Sensiscript RT Kit (Qiagen) and qPCR using QuantiTect SYBR Green PCR Kit (Qiagen) was employed for standard curve generation. The RT-PCR was performed in 20 μL volumes containing 2.0 μL 10 × buffer, 2.0 μL dNTP, 1.0 μL each primer, 0.2 μL RNase inhibitor, 1 μL RT, 10.8 μL RNase-free H2O, and 2 μL RNA, and amplified at 37°C for 60 min. The qPCR was performed in 25 μL volumes containing 12.5 μL of 2 × SYBR Green Mix, 1.5 μL each primer, 4 μL cDNA, and 5.5 μL H2O, using ABI 7900HT system (Applied Biosystems). The amplification cycle was as follows: 95°C 15 min, 40 cycles of 94°C 15 sec, 60°C (for MNV) or 56°C (for HAV) 30 sec, and 72°C 30 sec, followed by a dissociation step of 95°C 15 sec, 60°C 15 sec, and 95°C 15 sec. A standard curve was generated for each type of manure/biosolids containing HAV or MNV. The primer for MNV was MNV F: 5′-TCTTCGCAAGACACGCCAATTTCAG-3′, and MNV R: 5′-GCATCACAATGTCAGGGTCAACTC-3′, targeting the polymerase gene of MNV (Hsu et al., 2005). This primer pair produces a 318-bp product from nucleotides at positions 4228 to 4545. The HAV primer set targeted VP1 to VP3 capsid protein and produced a 192-bp product from nucleotides at positions 2035 to 2226 (Sair et al., 2002). The sequences are HAV3: 5′-CTCCAGAATCATCTCCAAC-3′, and HAV5: 5′- CAGCACATCAGAAAGGTGAG-3′.
Statistical analysis
Statistical analysis was performed with Tukey's test using JMP8 software to assess the data variations. The statistical significance was considered when p < 0.05. All samples were set in duplicate, and each experimental replicate was assayed in two replicates for both RT-qPCR and cell culture assay.
Results
MNV survival in manure/biosolids
For MNV stored in manure or biosolids at 20°C, viral genome was stable in RPL and SM, and after 60 days of incubation, there was no significant difference from viral RNA of day 0 (Fig. 1A). For MNV in AL, viral RNA concentration at day 60 was significantly lower than day 0, but still had >6 log RT-qPCRU/g (Fig. 1A). MNV genome degraded relatively quickly in PL and biosolids type 1, and after 60 days, there was significant loss of MNV RNA compared to day 0, with approximately 1.5–2 log reduction. Viral RNA was most unstable in liquid DM, and for storage at both 20°C and 4°C, the RNA was below the detection limit (∼3.7 log RT-qPCRU/mL) at 30 days. At 50 days, no viral RNA was detected in nondiluted DM. For other solids incubated at 4°C, MNV genome was quite stable and no significant loss of viral RNA was detected after 60 days of incubation (Fig. 1B) with the exception of MNV stored in biosolids type 1. No viral RNA of MNV was detected in either of the two high pH biosolids types 2 and 3 at time zero, indicating that viruses were degraded very quickly in high pH biosolids. For the infectivity study, the infectious concentration of MNV in PL, AL, RPL, and SM had no significant differences at either 20°C or 4°C after 60 days of incubation (Fig. 2A, B). In the liquid DM, the infectious MNV was below the detection limit (∼1000 PFU/mL) at 20 days at 20°C and at 30 days at 4°C (Fig. 2A, B). For both the MNV genome and infectivity, MNV incubated in manure or biosolids had no significant difference between 20°C and 4°C after 60 days. No infectious data were obtained for biosolids type 1 sample, as there was a strong cytotoxic effect on RAW 264.7 cells even at the 10−5 dilution (detection limit >5.7 log PFU/g). Other manure samples had cytotoxic effect on RAW 264.7 cells at 10−1 and 10−2 dilution.
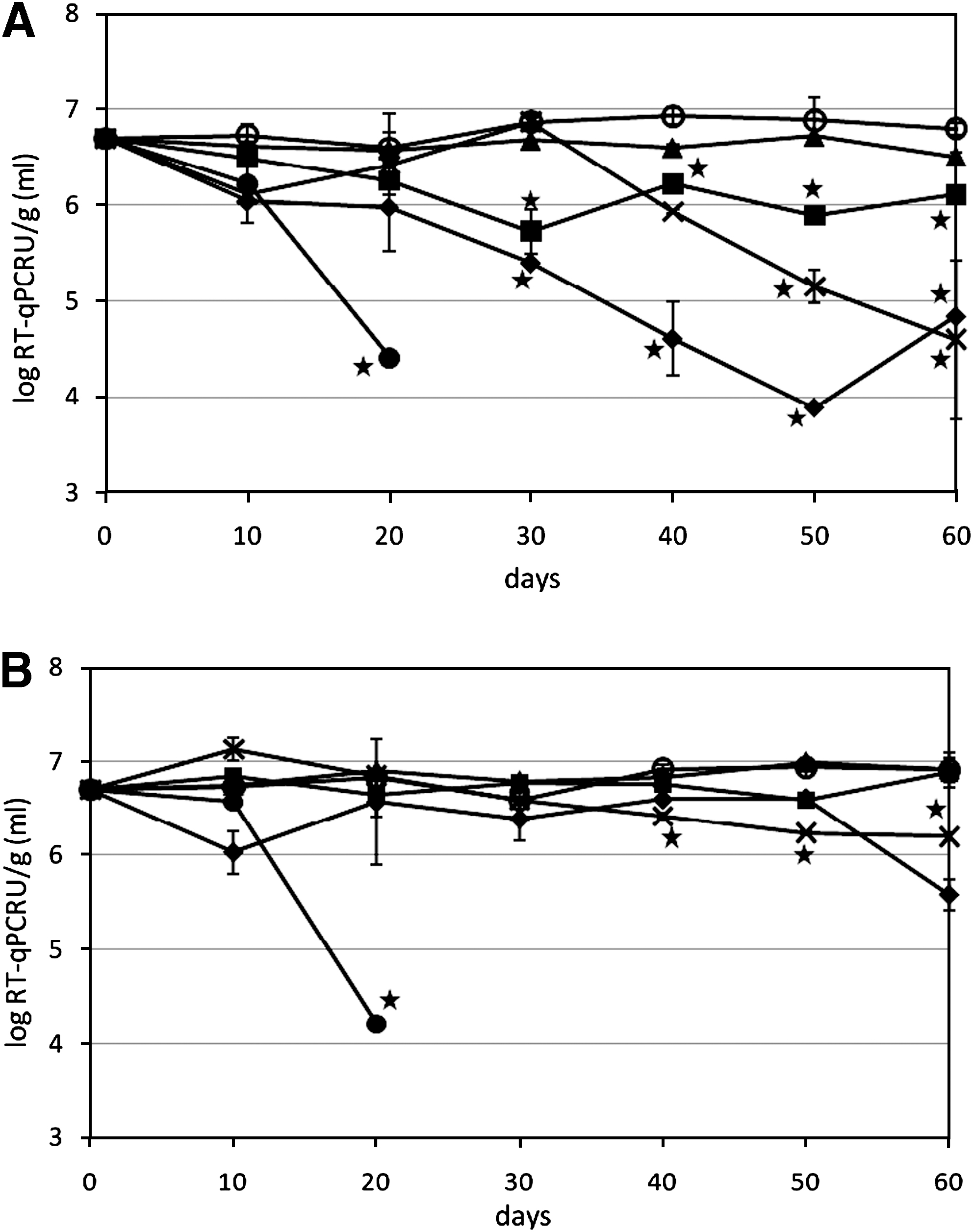
Survival of murine norovirus-1 viral genomes in manure and biosolids during 60 days of storage at (
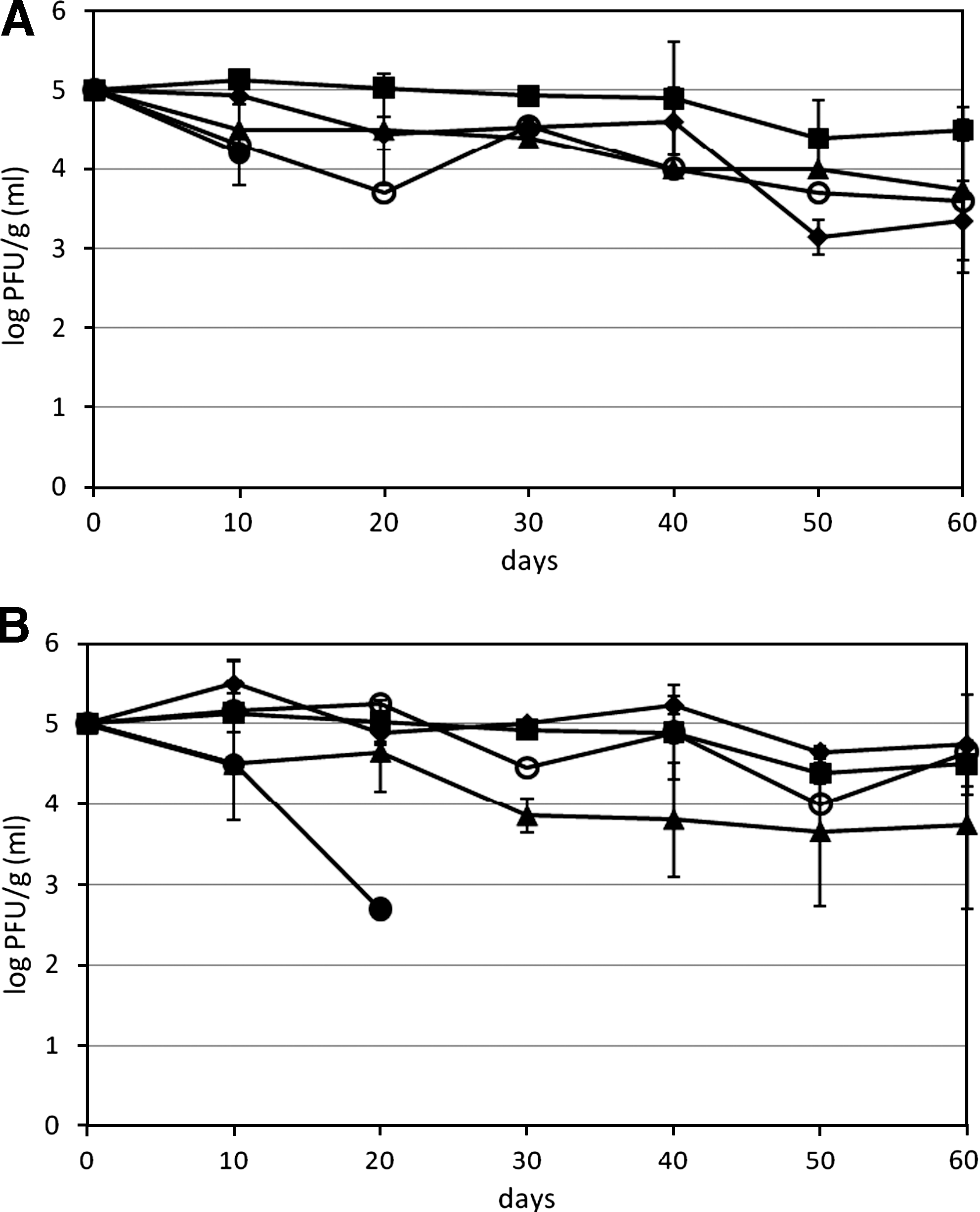
Survival of infectious murine norovirus-1 in manure and biosolids during 60 days of storage at (
HAV survival in manure/biosolids
For HAV at 20°C, significant loss of HAV RNA was observed in samples of biosolids type 1 and DM after 60 days, with ∼1–1.5 log reduction, whereas HAV RNA was stable in other manure (Fig. 3A). At 4°C, no significant loss of viral RNA was observed for each sample after 60 days of storage (Fig. 3B). When 20°C is compared with 4°C, HAV incubated in DM and biosolids type 1 had significantly higher viral RNA at 4°C than at 20°C after 60 days, whereas temperature had no significant effect on HAV genome in any other manure. Identical to MNV, no HAV RNA was detected from the high pH biosolids types 2 and 3 at time zero. For infectivity at 20°C, HAV in SM and RPL had significantly higher amounts of infectious virus than PL and AL after 60 days of incubation, and the inactivation of HAV in AL occurred as soon as 10 days (Fig. 4A). At 4°C, no significant differences were observed between PL and RPL, but HAV in AL had significantly lower numbers of infectious virus than HAV in RPL and SM. Concerning the temperature effect, HAV incubated in SM had significantly higher infectious virus at 4°C than at 20°C after 60 days, whereas HAV in PL, RPL, and AL were similar at both temperatures. For biosolids type 1, there were ∼ 4 log TCID50/g infectious HAV at 20 and 30 days at 20°C and 4°C, respectively, and after which, the titer decreased below the detection limit (∼3.8 log TCID50/g). The HAV was least infective after incubation in liquid DM, the infectious titer was below the detection limit (∼3.1 log TCID50/mL) at 10 days at 20°C, and at 4°C, there was 3.8 log at day 10, which decreased below the detection limit at 20 days. Biosolids type 1 suspension had cytotoxic effect on FRhK-4 cells at 10−2 dilution, whereas other manure suspensions had a cytotoxic effect at 10−1 dilution.
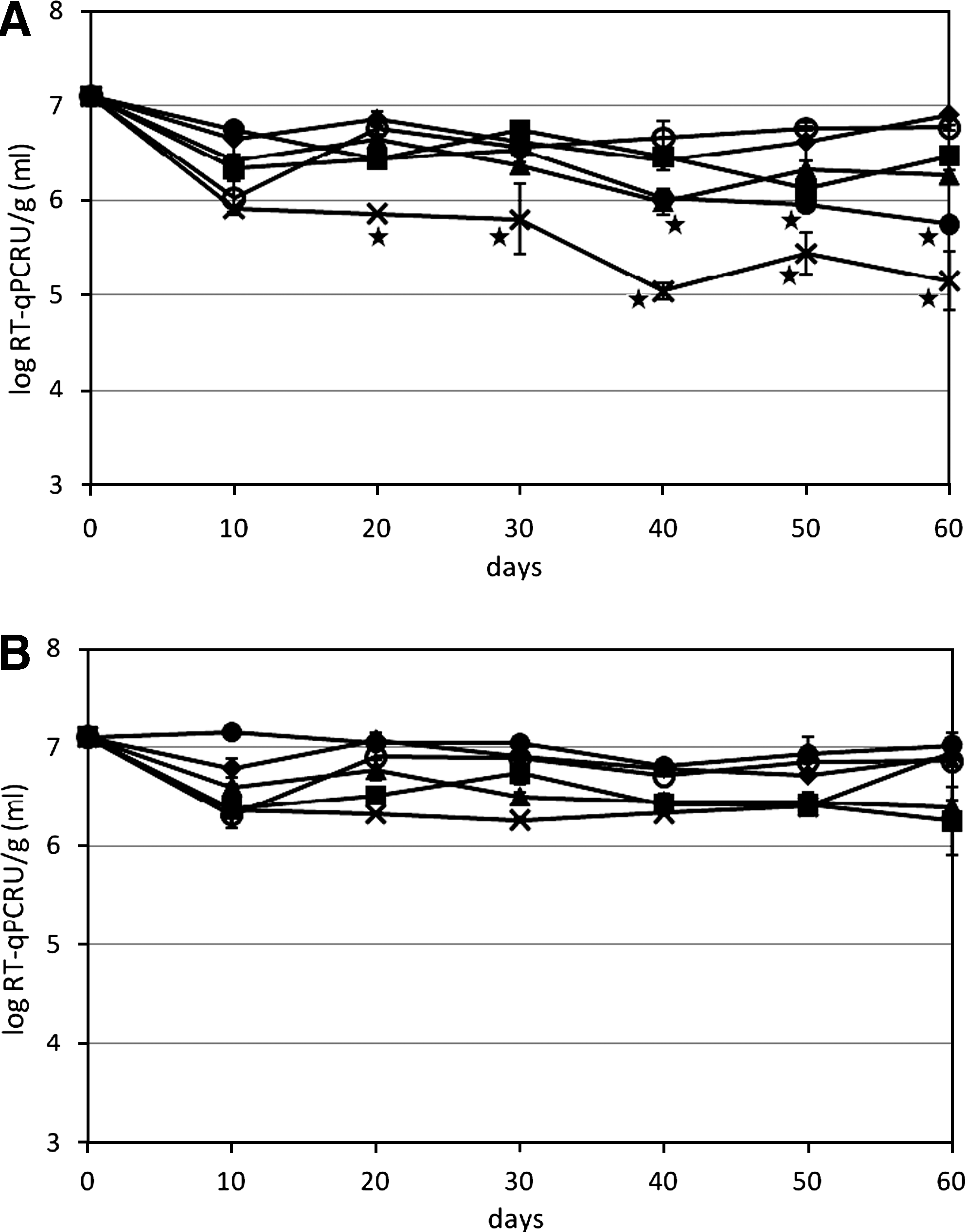
Survival of HAV viral genomes in manure and biosolids during 60 days of storage at (
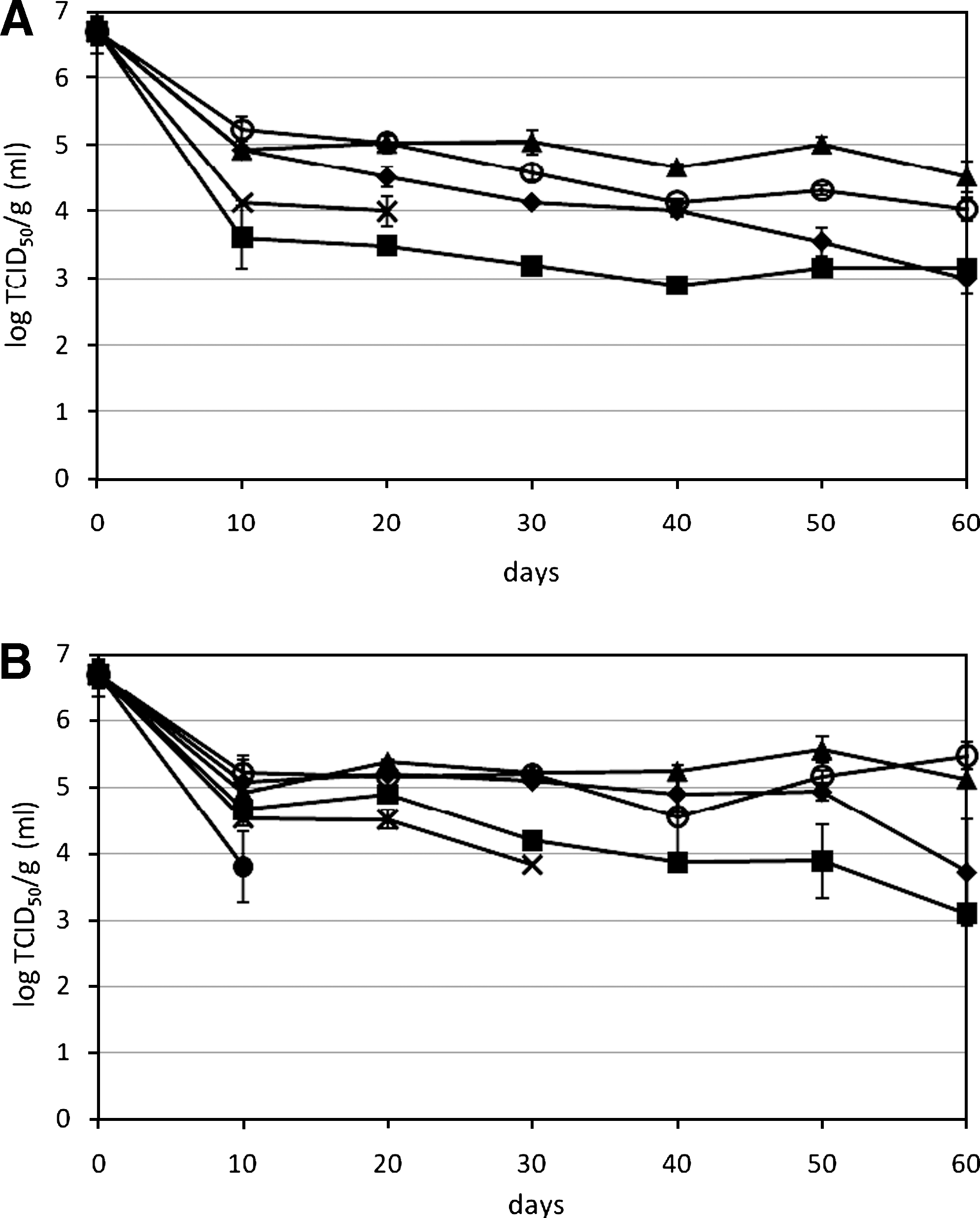
Survival of infectious HAV in manure and biosolids during 60 days of storage at (
Discussion
Although there is an increasing concern considering contamination of fresh produce on the field with human pathogens by application of manure/biosolids on agricultural land, few studies have been conducted on the survival of viruses in the manure and biosolids or soil environment. Pourcher et al. (2007) reported that after they inoculated ∼5.8 log genomic copies/g enteroviruses into WWTP sludge and applied onto soil, no virus was detected after 2 weeks during mid fall. However, after being spiked with anaerobic digestion residue and applied to soil, porcine parvovirus was persistent and no loss was obtained after 50 days at 15°C–18°C (Johansson et al., 2005). This study showed that MNV and HAV could maintain infectivity in animal manure after incubation for 60 days at both 20°C and 4°C. However, infectivity is virus and manure type dependent: the alum added in the AL sample seemed to have no effect on MNV, but did affect HAV infectivity and could inactivate some HAV at both 20°C and 4°C. Alum is used as a coagulant in drinking water treatment. It can destabilize and aggregate small particles such as bacteria and viruses (Matsui et al., 2003). It was reported that bacteriophages MS2 and f2 were inactivated when in contact with Al3+ in water (Yamamoto et al., 1964). In addition, after being added to poultry litter, aluminum hydroxide is formed that can absorb virus particles, and this strong absorption may result in virus inactivation (Jin and Flurry, 2002). It is not known why alum only had an effect on HAV; however, the inactivation of virus by metal and metal oxides was virus dependent. For example, iron oxides were found to absorb and inactivate MS2, but had no effect on phage φX174 (Chu et al., 2000). It is unknown why both viruses were unstable in DM, but the inactivation mechanisms on HAV and MNV may be different, as HAV RNA was stable but lost infectivity quickly, while the MNV genome was degraded. Overall, for both MNV and HAV, there were more genomic copies than infectious virus after storage for 60 days, showing that some virus particles were still present but lost infectivity, which could be due to loss of the virus' ability to attach to cell receptors resulting from capsid modifications (Nuanualsuwan and Cliver, 2003).
Temperature is the primary factor that affects virus survival in the soil, and virus inactivation rate is higher at high temperatures than at lower ones (Jin and Flurry, 2002). Under high temperature, virus may absorb more strongly to soil particles and are more likely to undergo structural changes that may cause the loss of infectivity. In addition, microbial activity in soil or manure increased with an increase in temperature. Deng and Cliver (1995) showed that 1.5–2.5 log virus particles/mL reduction of HAV was achieved after incubation in a mixture of septic tank effluent and swine or dairy cattle manure slurry for 70 days at 5°C; however, at 22°C, there was 4–5 log virus particles/mL loss after 70 days. However, in this study, the infectivity of HAV had no significant difference between 20°C and 4°C in three types of poultry litter, but HAV in biosolids type 1, SM, and DM maintained significantly higher infectivity at 4°C than 20°C. The inconsistency concerning temperature effects on HAV compared to the previous study may be due to the lack of microbial activity in all the manure samples.
Microbial activity is one of the antiviral properties of manure and biosolids. However, current literature is inconsistent regarding the effect of autochthonous microorganisms on virus inactivation. It has been reported that inactivation of virus in nonsterile soil was greater than sterile soil (Sobsey et al., 1980; Hurst, 1988; Nasser et al., 2002; Davies et al., 2006). However, others showed that higher inactivation was observed in sterile soil or ground water (Alvarez et al., 2000; Ottosson and Stenström, 2003; Zhao et al., 2008), while John and Rose (2005) found no clear difference on virus inactivation between nonsterile and sterile conditions in ground water. The effect of sterilization on virus is also soil type dependent, and increasing inactivation may be achieved with soil containing high metal oxides due to the strong absorption of virus to soil particles after sterilization (Zhao et al., 2008). The manure and biosolids used in this study were autoclaved to facilitate cell culture studies used in this work.
The treatment of biosolids also affected virus survival. The immediate inactivation of MNV and HAV in biosolids types 2 and 3 was likely due to the high pH of these two lime-treated biosolids. It was reported that fecal coliform bacteria and reovirus were completely inactivated in lime stabilized biosolids (Abu-orf et al., 2004) and likewise in research conducted by Wei et al. (2009) using these same biosolids types 2 and 3, human adenovirus 41 lost infectivity immediately after inoculation. For biosolids type 1, FeCl3 was added for phosphate control and the iron content was significantly higher than that of the other two types of biosolids (Table 1), SM and DM (Shober and Sims, 2009). The presence of iron oxides was shown to improve the absorption and inactivation of MS2 (Chu et al., 2000). As shown in the study, both HAV and MNV RNA were unstable in biosolids type 1 at 20°C, but were rapidly inactivated in alkaline pH.
Conclusion
Both HAV and MNV can maintain some infectivity in manure and type 1 biosolids after storage for up to 60 days at both 20°C and 4°C. Both viruses were inactivated most rapidly in lime-stabilized biosolids with alkaline pH and were unstable in liquid DM, but alum added in poultry litter had different effects on the two viruses. Alum inactivated some HAV at both temperatures but had no effect on MNV. This indicates that application of soil amendments to agriculture lands may pose a risk to food safety and human health. However, virus stability and infectivity is dependent on virus, manure, and biosolids type.
Footnotes
Acknowledgment
This project was funded in part by the USDA National Research Initiative Watershed Grant no. 2006-35102-17405. The authors thank Dr. Alyssa Collins for her work in obtaining the biosolids and animal manure.
Disclosure Statement
No competing financial interests exist.