
Abstract
In a previous study we showed that an Escherichia coli O157:H7 strain that was unable to form biofilm was retained in large numbers in dual-strain biofilms formed with an E. coli O-:H4 companion strain. In this study we tested additional companion strains for their ability to retain E. coli O157:H7 strain 0475s. Companion strains producing biofilm that withstood aggressive washes were able to significantly increase serotype O157:H7 retention. Dual-strain biofilms with certain companion strains retained higher percentages of strain 0475s, and that ability was independent of biofilm total cell numbers. Tests with additional non-biofilm-forming E. coli O157:H7 strains showed that enhancement by companion strains was not unique to strain 0475s. Experiments using an E. coli companion strain with deletions of various curli and cellulose genes indicated that dual-strain biofilm formation was dependent on companion strain properties. Strain 0475s was not able to generate biofilm or persist on plastic when grown in broth with a biofilm-forming companion and separated by a 0.2 μm porous membrane, indicating a requirement for intimate contact with the companion strain. When dual-strain biofilms and planktonic cells were challenged with 5% H2O2, strain 0475 showed greater survival in biofilms with certain companion strains compared to the corresponding planktonic cells. The results of this study indicate that non-biofilm-forming E. coli O157:H7 strains are retained on solid surfaces associated with biofilms generated by companion strains. However, properties other than biofilm mass enable certain companion strains to retain greater numbers of E. coli O157:H7.
Introduction
T
In some studies, Escherichia coli O157:H7 has been shown to attach and persist on solid materials found in meat-processing plants (Stopforth et al., 2003; Skandamis et al., 2009). Other studies have shown that strains of O157:H7, grown in a single-strain culture, do not form robust biofilm on solid surfaces, but rather form small clusters of attached cells (Dewanti and Wong, 1995; Uhlich et al., 2006, 2008) or completely fail to attach (Klayman et al., 2009). A major mechanism for biofilm formation in E. coli and many of the Enterobacteriaceae involves expression of the adhesin, curli fimbriae, and/or the exopolysaccharide, cellulose (Zogaj et al., 2003; Römling, 2005). Strains producing curli fimbriae exhibit a brown, dry, and rough (bdar) colony morphology on agar containing Congo red dye, whereas strains producing cellulose produce a pink, dry, and rough (pdar) morphology (Zogaj et al., 2001). Strains expressing both cellulose and curli have a red, dry, and rough (rdar) phenotype, and colonies of strains expressing neither cellulose nor curli appear smooth and white (saw) (Zogaj et al., 2001). From 46% to 79% of commensal E. coli isolates were found to produce curli, cellulose, or both depending on temperature (Bokranz et al., 2005). Those strains expressing either one or both components had medium to high biofilm-forming capabilities.
A number of recent studies have focused on the ability of different species of bacteria to retain serotype O157:H7 in mixed-species biofilms or increase its adhesion to solid surfaces. Pseudomonas aeruginosa retained E. coli O157:H7 in a mixed-species biofilm formed on glass (Klayman et al., 2009). A variety of bacterial isolates from a meat-processing environment increased the attachment of E. coli O157:H7 to polyurethane when cocultured at 15°C (Marouani-Gadri et al., 2009). In a study of isolates from two multistate outbreaks of E. coli O157:H7, a serotype O157:H7 strain unable to form biofilm in pure culture persisted on glass in a dual-strain biofilm generated together with a different E. coli serotype isolated from the same outbreak (Uhlich et al., 2008). These findings suggest that non-biofilm-forming E. coli O157:H7 strains may be retained in food production environments through interactions with other contaminating bacteria. The aim of the current study was to investigate the retention of E. coli O157:H7 on glass when cultured together with different serotypes of biofilm-forming E. coli, other members of Enterobacteriaceae, and strains of Pseudomonas and Bacillus.
Materials and Methods
Bacterial strains, media, and antibiotics
E. coli strains 06E01767 (serotype O-:H4) and 06F00475 (serotype O157:H7) were isolated from the 2006 E. coli O157:H7 multistate outbreak associated with spinach (Uhlich et al., 2008). Strain 0475s is a streptomycin-resistant strain made by passage of strain 06F00475 on brain heart infusion (BHI; Becton Dickinson, Sparks, MD) agar containing increasing streptomycin concentrations. E. coli of bovine origin (Table 1) were isolated at the Kansas State University Veterinary Diagnostic Laboratory (Manhattan, KS) and serotyped at the E. coli Reference Center at The Pennsylvania State University (University Park, PA). Rifampicin-resistant serotype O157:H7 strains 9, 20, 21, and SAL1 were from the Microbial Food Safety culture collection (Eastern Regional Research Center, Wyndmoor, PA). Strains 9 and SAL1 contain the Shiga toxin 1 (Stx1 ) gene, whereas strains 20 and 21 contain both Stx1 and Stx2 genes. Other strains in this study are described in Table 1. Strains were propagated on BHI agar. Luria–Bertani (LB; Becton Dickinson) plates with calcofluor, used for detection of cellulose production, and Congo red indicator agar were prepared as described (Hammar et al., 1996; Uhlich et al., 2006). Streptomycin and rifampicin were added to agar plates as needed at concentrations of 100 and 50 μg/mL, respectively.
Formed biofilm on glass that was detached during washes.
Formed biofilm on glass and medium surface that was detached during washes.
−, Not visible; +, thin zone; ++, intermediate zone; +++, wide zone;
Construction of deletion mutant strains
Strains of 06E01767 with deletion of the csgA (strain 1767-CsgA), csgB (strain 1767-CsgB), csgD (strain 1767-CsgD), bcsC (strain 1767-BcsC), and both the csgA/bcsC (strain 1767-A/C) genes were constructed using the Quick and Easy Gene Deletion Kit (Gene BridgesGmbH, Heidelberg, Germany) as previously described (Uhlich, 2009). Forward and reverse primers were designed from the EDL933 genome sequence (Accession no. AE005174) to provide 60-bp of upstream and downstream sequence homology that terminated with the start and stop codons of the target genes. Polymerase chain reaction (PCR) with kit template produced linear product consisting of the target genes and flanking sequences where the entire sequence between start and stop codons was replaced by a neomycin resistance cassette. Linear PCR products with the mutated genes were integrated in strains by Red/ET recombination, and the neomycin cassettes were resolved from all chromosomal integrations generating nonpolar mutations by the manufacturer's protocols (Gene BridgesGmbH). Gene deletions were verified by PCR using primers (not shown) that were designed from sequences outside of the chromosomal regions that provided homology for recombination. The wild-type and mutant strains achieved similar maximum population densities after 72-h incubation at 25°C in LB broth without added salt (LB-NS) (results not shown).
Biofilm assays
Glass microscope slides (3 in. × 1 in. × 1 mm) provided a large supply of uniform coupons and were used in all biofilm studies to reduce experimental variability. Mixed-strain biofilm assays were performed as previously described (Uhlich et al., 2008) with the following modifications. E. coli O157:H7 strains and companion strains were differentiated by their antibiotic resistance. Starter cultures and biofilms were generated in LB-NS broth and containing no antibiotics. Biofilms were washed aggressively with 25 mL phosphate-buffered saline (PBS) by vortexing thrice for 30 s on a Daigger Vortex Genie 2 at setting 8 (Daigger, Vernon Hills, IL). Detached biofilm cells were spin-plated on BHI agar (total count) or BHI agar containing the appropriate antibiotic (serotype O157:H7 count) to enumerate and differentiate cell populations. For biofilm observation, slides or tissue culture wells and inserts were stained with 0.1% crystal violet for 20 min at 25°C. Biofilm removal from slides was verified by loss of staining. P. aeruginosa, Citrobacter freundi, Citrobacter braakii, Serratia liquefaciens, and Bacillus subtilis generated wider zones of biofilm at the air–media interface in tryptic soy broth (TSB; Becton Dickinson) than in LB-NS, or 1/20th strength TSB (results not shown). Therefore, TSB was substituted for LB-NS in biofilm experiments with those organisms.
Dual-strain biofilm studies with physical separation of strains were performed in six-well, nontreated, polystyrene multidishes fitted with 25-mm tissue culture inserts containing 0.2 μm anapore membranes (Nunc, Roskilde, Denmark). Each chamber was loaded with 4 mL of LB-NS inoculated with 40 μL of either strain 06E01767 (inner chamber) or strain 0475s (outer chamber). Dishes were incubated at 25°C for 72 h. For enumeration of strain 0475s, inserts were removed and multidish wells were rinsed thrice with 8 mL PBS by sealing with parafilm and vortexing for 30 sec. Cells were dislodged from the wells into 8 mL of sterile 0.1% peptone water by scraping with a sterile spatula and vortexing with 0.3 g glass beads. Cells were enumerated by dilution and spread-plating on BHI containing 100 μg/mL streptomycin.
Peroxide resistance studies
The resistance of biofilm bacteria to 5% H2O2 was determined as previously described (Uhlich et al., 2006). Briefly, mixed-strain biofilms were generated and washed as described above. The biofilms were challenged with 5% H2O2 for 10 min and washed once with 1% sodium pyruvate to neutralize the H2O2 and once with PBS. The total surviving bacteria and antibiotic-resistant serotype O157:H7 were enumerated by dilution and spread-plating. Planktonic cell samples were harvested from the supernatant of 72-h biofilm tubes and challenged with H2O2 as described (Uhlich et al., 2006), and cell populations were enumerated by dilution and spread-plating.
Statistical analyses
Comparisons of total bacteria and E. coli O157:H7 mean cell counts recovered from biofilm and planktonic samples were by analysis of variance, and the significance of the means was determined by the Bonferroni technique (Miller, 1981). Logarithmic transforms of the percentages of E. coli O157:H7 in either dual-strain biofilms or dual-strain broth cultures were estimated by taking the difference of the log10 colony forming units (CFU)/mL values for E. coli O157:H7 and total cell counts. These were subjected to analysis of variance and Bonferroni separation.
Results and Discussion
E. coli O157:H7 strain 0475s retention on glass is enhanced by biofilm formation with companion strain 06E01767
Strain 06E01767 (O-:H4) was recovered from single-culture biofilms in >6 log10 CFU/mL, whereas recovered strain 0475s (O157:H7) counts (0.33 log10 CFU/mL) were not significantly different from 0 (p > 0.05) (Table 2). However, in a dual-strain biofilm, strain 0475s was recovered in >4 log10 CFU/mL, confirming earlier results where 06E01767 greatly enhanced O157:H7 retention (Table 2) (Uhlich et al., 2008). Unlike the earlier study, the O157:H7 strain in this study was recovered in fewer numbers than strain O-:H4. This difference appeared to be due to the method used for strain differentiation. In the previous study, strains were differentiated by colony fluorescence after transformation with either plasmid pUC19 or pUC19 bearing green fluorescent protein (GFP). In the current study, chromosomal antibiotic resistance was used for strain differentiation to avoid potential problems with GFP intolerance (Galen et al., 1999). Additional experiments (results not shown) demonstrated that strain O-:H4 carrying either pUC19 or pUC19-GFP attained a 2 log10 CFU/mL lower maximum population density in BHI broth compared to the wild-type strain. Maximum cell densities of E. coli K-12 or O157:H7 were unaffected by either plasmid. It is unclear why pUC19 is unstable in strain 06E01767, but it has been shown that rho mutations in E. coli can lead to pUC19 instability (Sozhamannan et al., 1999). Additional studies are needed to determine how pUC19 affects strain 06E01767 survival. However, it is clear that results using plasmid marker systems for differentiation of strains in growth-dependent assays must be interpreted with caution.
Indicates counts of strain 0475s recovered from dual-strain biofilms that were not significantly different (p > 0.05) from counts of strain 0475s recovered from single-strain biofilm experiments.
Formed biofilm on glass that was detached during washes.
Formed biofilm on glass and medium surface that was detached during washes.
−, Not visible; +, thin zone; ++, intermediate zone; +++, wide zone; CFU, colony forming units; SD, standard deviation.
E. coli strain 0475s retention on glass is enhanced by biofilm-forming companion strains that form biofilm
In comparisons of other Enterobacteriaceae companion strains for their ability to retain serotype O157:H7, we used fresh, minimum-passage (3 or less passages) bovine isolates of E. coli to avoid lab adaptation, which may attenuate biofilm formation (Römling, 2005). All E. coli, except strain M15, that generated visible biofilm retained from >2 log10 CFU/mL to nearly 5 log10 CFU/mL of 0475s cells (Table 2). These results indicate that biofilm-producing companion strains other than 06E01767 can retain strain 0475s. Strains with the highest cell counts did not necessarily contain the highest 0475s percentages. Both strain M1 and M11 biofilms had total counts >6 log10 CFU/mL but contained <0.1% strain 0475s cells. It is also clear that strain 0475s is retained better in biofilms formed with certain companion strains, and this increased retention is not dependent on biofilm mass or cell numbers. All E. coli strains that failed to generate visible biofilm were associated with 0475s counts below detectable levels.
We also tested E. coli O26:H11 strain H30 that does not bind Congo red and may use a different mechanism compared to curli or cellulose expression for biofilm formation (Fratamico et al., 2009). Strain H30 generated visible biofilm containing >6 log10 CFU/mL bacteria. Like the Congo red–binding E. coli, H30 also enhanced strain 0475s numbers on glass, indicating that strains using different mechanisms for biofilm formation can also retain strain 0475s.
Salmonella strains H3502 (Salmonella Enteritidis), 02-216 (Salmonella Newport), and H1275 (Salmonella Newport) all increased strain 0475s recovery compared to 0475s in single-strain biofilms (Table 2). Strain 35551 (S. liquefaciens) failed to form visible biofilm, although nearly 5 log10 CFU/mL of total cells were recovered after the washes. However, strain 0475s was not recovered. Citrobacter strains formed substantial biofilm that remained after washes but retained some of the lowest percentages of strain 0475s in the study. These results suggest possible genus differences among the Enterobacteriaceae in their ability to retain O157:H7, and confirm that companion strains differ in their ability to enhance strain 0475s retetion on glass.
B. subtilis and P. aeruginosa did not increase the recovery of E. coli O157:H7 (Table 2). B. subtilis SMS18 formed a biofilm on glass and spread across the medium surface, but the washes removed all visible material. B. subtilis cells were recovered from the slides in >3 log10 CFU/mL after the washes, but the 0475s counts were not significantly different from 0 (p > 0.05). Likewise, P. aeruginosa PG201 formed biofilm that was removed by aggressive washes. Some P. aeruginosa were recovered, but 0475s counts were not significantly different from 0 (p > 0.05). Other studies found that Pseudomonas did not persist well in food plants, although Bacillus did, and both increased the attachment of E. coli O157:H7 to polyurethane (Marouani-Gadri et al., 2009). E. coli O157:H7 has been reported to persist in biofilms formed with P. aeruginosa but not on glass when cultured as a single strain in flow cell studies (Klayman et al., 2009). In the current study, no E. coli O157:H7 cells were recovered after aggressive washes of the PG201 or SMS18 biofilms. We did not determine the cell populations in the Pseudomonas or Bacillus biofilms that were dislodged by washing, and strain 0475s could have been present in high numbers in the detached biofilms. However, since substantial numbers of attached Pseudomonas and Bacillus strains were still recovered after the washes and no associated O157:H7 was detected, it may be that the 0475s attachments were weaker, allowing strain O157:H7 removal. Alternatively, O157:H7 cells may have been more associated with the upper levels of the biofilm and not attached to the glass surface.
The retention of serotype O157:H7 strains other than 0475s is also enhanced by dual-strain biofilm formation
Additional strains of E. coli O157:H7 were also tested for their ability to be retained in dual-strain biofilms with various companion strains (Table 3). Companion strains M11, M17, M23, and M25 were each tested with O157:H7 strains 9, 20, 21, and SAL1. The serotype O157:H7 strains tested as single strains formed no visible biofilm on glass, and recovered cells ranged from 1.06 to 1.35 log10 CFU/mL (Table 3). In dual-strain biofilms, recovered E. coli O157:H7 counts ranged from >3 to >5 log10 CFU/mL, similar to strain 0475s numbers retained in biofilms with the same companion strains. This demonstrates that O157:H7 strains other than 0475s are also retained in greater numbers on glass when accompanied by biofilm-forming companion strains. As with strain 0475s, retention of each rifampicin-resistant strain was significantly higher (p < 0.05) in combination with E. coli strains M23 and M25 than with strains M11 and M17.
The results of Bonferroni means separation for comparisons of the quantity of serotype O157:H7 present in mixed-strain biofilms are shown using lower-case letters in parentheses, where values with the same letter are not significantly different (p > 0.05) from each other.
Competition between strains during dual-strain biofilm formation does not fully determine the level of E. coli O157:H7 retention within the biofilm
The low retention of serotype O157:H7 in certain dual-strain biofilms could result from an inhibitory or lethal affect of a companion strain on planktonic E. coli O157:H7 cells during biofilm development. To investigate this possibility we determined the cell populations of planktonic cells for each O157:H7–companion strain pair. The percentages of serotype O157:H7 comprising the 72-h biofilms and the surrounding planktonic cells are shown in Table 3. In general, the percentage of O157:H7 in both biofilms and planktonic cells was less than 50%, indicating that most companion strains maintained a higher population density than serotype O157:H7 in mixed cultures. However, none of the companion strains completely inhibited strain O157:H7 growth. Moreover, the dual-strain pairs forming biofilms with the lowest percentages of serotype O157:H7 (M11/20, M17/20, M11/21, and M17/21) did not have the lowest percentages of O157:H7 in the surrounding broth. These results indicate that the low percentages of serotype O157:H7 retained by certain companion strains in dual-strain biofilms were not due to inhibition of O157:H7 growth in the planktonic phase. It should also be noted that for pairs M23/9 and M23/SAL1, the percentages of E. coli O157:H7 cells in biofilms were greater than the percentages of planktonic cells, indicating that strain M23 may recruit certain O157:H7 strains into biofilms in greater numbers.
Dual-strain biofilm formation depends on the biofilm properties of the companion strain
Although E. coli O157:H7 was retained in dual-strain biofilms, it is unclear whether it participated in biofilm formation or simply colonized the companion strain biofilm. Most companion strains in this study showed Congo red binding or calcofluor staining indicative of curli and cellulose expression (Table 1). We selectively deleted certain curli and cellulose genes in the rdar strain 06E01767 to determine their effect on dual-strain biofilm formation. Curli expression is mediated through the csgDEFG and csgBAC operons. CsgA is the curli subunit protein, CsgB is a nucleator protein, and CsgD is a transcriptional regulator of both curli and cellulose synthesis (Römling, 2005). The csgE, csgF, and csgG genes are involved in curli export (Robinson et al., 2006). CsgA is secreted from the cell and polymerizes on the surface of cells displaying CsgB. Strains deficient in CsgA, but expressing CsgB, can polymerize curli through interbacterial complementation if a CsgA-expressing strain is in proximity. The bcsABZC operon encodes the structural genes required for cellulose synthesis. BcsA and BcsB form the cellulose synthase complex, BcsZ is a cellulase, and bcsC encodes a putative oxidase required for cellulose synthesis (Römling, 2002). Nonpolar deletion of csgD or csgA and bscC eliminated biofilm formation and generated a saw phenotype in strain 06E01767 (Table 1). Deletion of only csgA or csgB produced a pdar phenotype, whereas bcsC deletion produced bdar colonies. All three (csgA-, csgB-, and bcsC-deleted) mutant strains formed biofilm, but these were attenuated compared to that of strain 06E01767 (Table 1).
The results of dual-strain biofilm comparisons using the mutant strains are shown in Figure 1. Strain 0475s paired with strains 1767-CsgD or 1767-A/C formed no visible biofilm, and the retained 0475s cells were not significantly different from 0 (p > 0.05), although 2.0 and 2.54 log10 CFU/mL of the respective companion cells remained adhered to the glass. Strains of 06E01767 deficient in csgA, csgB, or bcsC each formed slightly less biofilm than the wild-type strain, and each retained significantly less strain 0475s than the wild-type biofilm (p < 0.05), but significantly greater strain 0475s than biofilm generated with strains missing csgD or csgA/bcsA (p < 0.05). The reductions in serotype O157:H7 retention resulting from deletions of the various curli and cellulose genes were proportionate to the reductions in total biofilm, with no cells recovered when stainable biofilm architecture was completely lost. These results indicate the importance of curli and cellulose components for biofilm formation, and that dual-strain biofilm formation and O157:H7 retention within the biofilm are both dependent on the biofilm-forming properties of the companion strain.
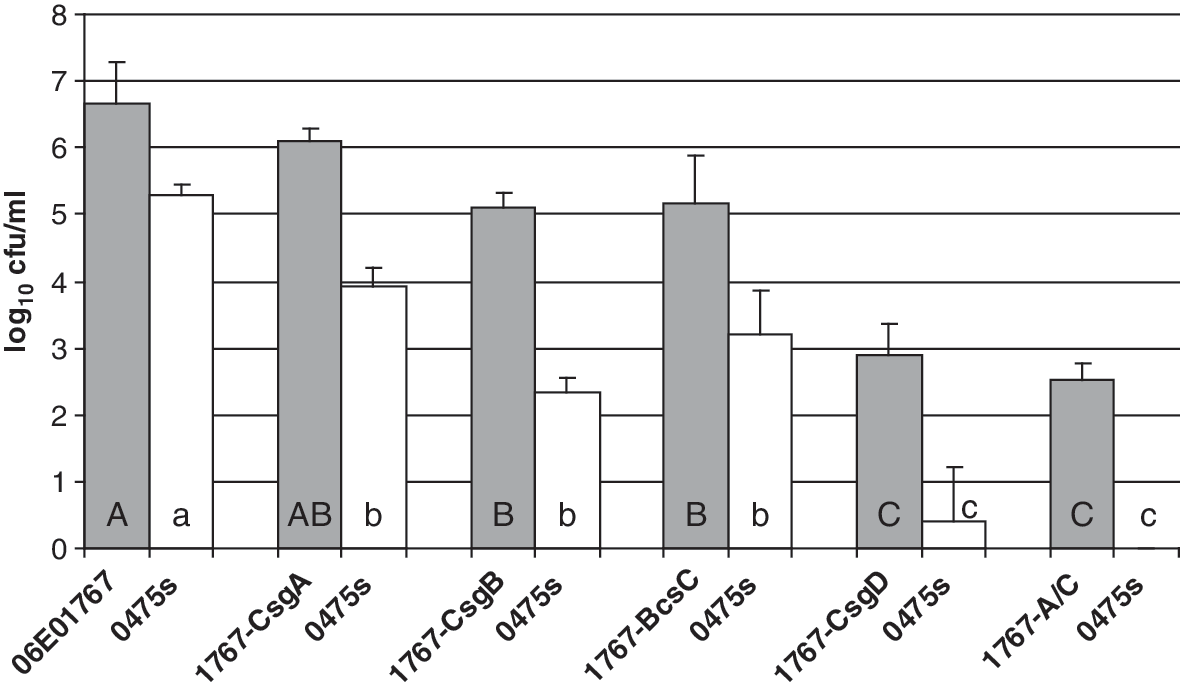
Mean log10 CFU/mL ± standard error of the total bacterial (shaded bars) and Escherichia coli strain 0475 (white bars) counts from dual-strain biofilms generated from combinations of strain 0475s with companion strains 06E01767, 1767-CsgA, 1767-CsgB, 1767-BcsC, 1767-CsgD, and 1767-A/C. Means were calculated from duplicate counts determined from four independent biofilm samples for each strain pair. Letters represent the results of a Bonferroni means separation for total bacterial counts (upper case) or strain 0475s counts (lower case), where counts with the same letter are not significantly different (p > 0.05) from each other.
Increased retention of E. coli O157:H7strains on glass requires intimate contact with companion strains
When strain 0475s was cultured together with strain 06E01767 in a common medium but physically separated by a 0.2 μm membrane, biofilm formed at the air–liquid interface on the strain 06E01767-side of the membrane (results not shown). There was no stainable biofilm on the strain 0475s-side of the membrane. When the adherent cells of the compartment enclosing strain 0475s were dislodged and enumerated, the counts (mean log10 CFU/mL = 3.37; standard error = 0.05) were not significantly different from those recovered from control wells where strain 0475s was separated from LB-NS medium without strain 06E01767 (mean log10 CFU/mL = 3.33; standard error = 0.17) (p > 0.05). The inability of strain 0475s to form biofilm when separated from the companion strain indicates a requirement for direct contact between the strains and argues against the initiation of independent biofilm formation by the O157:H7 strain through sharing of small soluble signaling factors or small individual biofilm components.
E. coli O157:H7 retained in certain dual-strain biofilms have increased resistance to the bacteriocidal effects of H2O2
Planktonic and biofilm cells from four different dual-strain pairs (strain 0475 with either companion strain M23, M25, 43162, or 8090) were challenged with H2O2 (Fig. 2). For each dual-strain pair, the numbers of total planktonic cells and 0475s planktonic cells surrounding each biofilm were significantly reduced after exposure to 5% H2O2 (p < 0.05). The total cell counts of each strain pair in unchallenged biofilms were approximately 6 log10 CFU/mL and the counts were not significantly different from each other (p > 0.05), indicating that total biofilm mass produced by dual-strain pairs was the same. Strain 0475s was retained in >3 log10 CFU/mL in each dual-strain biofilm except for strain 43162 (C. braakii), for which biofilm was undetectable. After exposure to H2O2, both the total cell counts and the strain 0475s counts from biofilms with strains M23 or M25 were significantly reduced compared to their respective untreated samples (p < 0.05). For the M23 dual-strain pair, significantly greater numbers of 0475s were recovered from the biofilm samples compared to the planktonic samples (p < 0.05). For the M25 dual strain pair, 0475s cells were recovered from the biofilm samples but not the planktonic samples. In strains 43162 and 8090 biofilms with strain 0475s, both the companion and O157:H7 strains were completely inactivated by 5% H2O2. These results indicate that serotype O157:H7 cells retained in certain dual-strain biofilms have increased H2O2 resistance compared to that of planktonic cells. The E. coli companion strains were both more resistant to H2O2 and more accomplished at retaining serotype O157:H7 than the Citrobacter companion strains. Therefore, the non-biofilm-forming serotype O157:H7 was able to survive H2O2 challenge when cocultured with E. coli but not with Citrobacter.
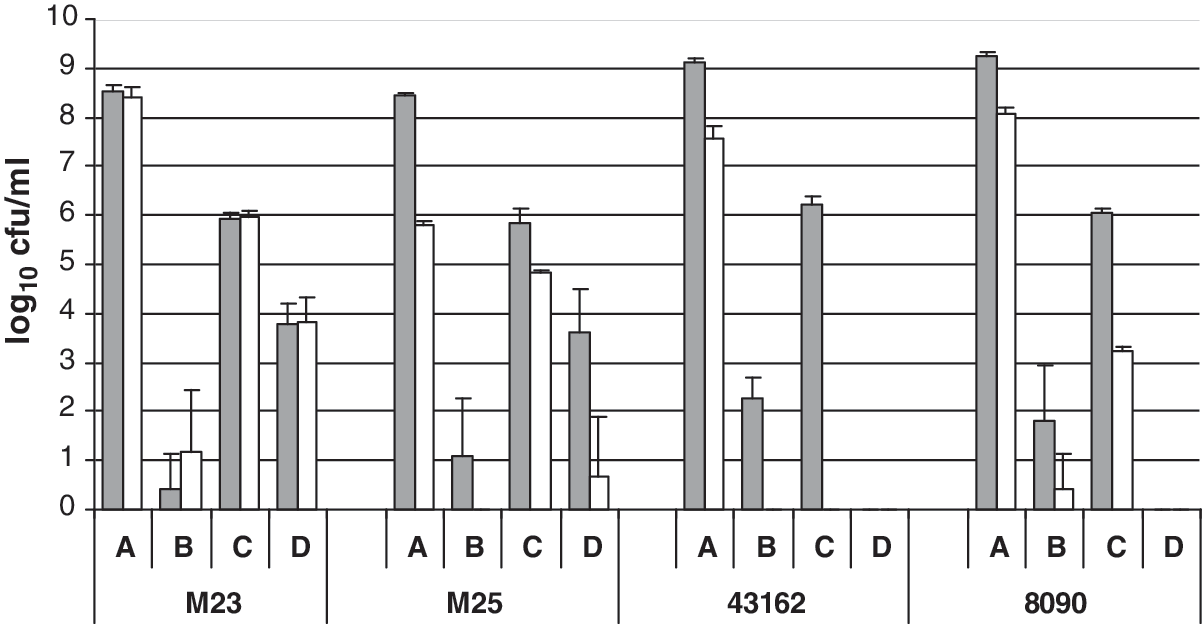
H2O2 challenge of dual-strain biofilms formed with E. coli O157:H7 strain 0475s paired with companion strains M23, M25, 43162, and 8090. The mean log10 CFU/mL ± standard error of the total cell counts (shaded bars) and the strain 0475s counts (white bars) recovered from (A) planktonic cells exposed to phosphate-buffered saline, (B) planktonic cells exposed to H2O2, (C) biofilm cells exposed to phosphate-buffered saline, or (D) biofilm cells exposed to H2O2 are shown. Means were calculated from duplicate counts determined from four independent biofilm samples for each strain pair–treatment combination.
We previously showed that an E. coli O157:H7 strain that overexpressed curli formed biofilm and had increased resistance to H2O2 compared to planktonic cells (Uhlich et al., 2006). Here, we showed that serotype O157:H7 strains that do not overexpress curli in dual-strain biofilms can also resist lethal doses of H2O2. The peroxide studies again demonstrated that different companion strains differ greatly in their ability to retain E. coli O157:H7. Although the total numbers of cells recovered from PBS-exposed dual-strain biofilms generated with strains 8090, 43162, M23, and M25 were not significantly different from each other (p < 0.05), the numbers of recovered strain 0475 cells ranged from 0 to almost 6 log10 CFU/mL. These results suggest that when E. coli O157:H7 is retained in mixed-strain biofilms with other contaminating bacteria, their resistance to antimicrobial agents such as H2O2 may depend more on the type of companion strain rather than the mass of the biofilm.
Conclusions
In this study we showed that although many strains of E. coli serotype O157:H7 do not form biofilm and are easily eliminated from surfaces by aggressive washes, their retention in dual-strain biofilms may provide an alternative mechanism for their persistence on solid surfaces. Different strains of Enterobacteriaceae were shown to be suitable companion strains for enhancing serotype O157:H7 retention on glass. The major requirements of these companion strains to enhance O157:H7 retention were that they form biofilm and have intimate contact with the serotype O157:H7 strain. It appears that E. coli O157:H7 does not form additional biofilm or contribute to existing biofilm in dual-strain cultures, but may simply colonize biofilm generated by the companion strain. It is unclear which portions of the biofilm are associated with O157:H7. Additional microscopic studies using fluorescent strains are needed to fully define the relationship between serotype O157:H7 and companion strains, and the location of serotype O157:H7 within the mixed-strain biofilm. Some companion strains were clearly more accomplished at allowing retention of serotype O157:H7 within their biofilm. These differences may be attributable to the companion strain, the biofilm matrix, or even the O157:H7 strain. In addition, these studies indicated that companion strains can also influence serotype O157:H7 resistance to H2O2. When studies of sanitizers and sanitization studies are conducted, multiple strains in coculture should be employed to simulate real world conditions.
Collectively, these data indicate that the persistence and peroxide resistance of E. coli O157:H7 in various environmental niches may be greatly influenced by both the resident microflora and the accompanying contaminating strains.
Footnotes
Acknowledgments
We thank Bryan Cottrell and Jennifer Elmer for technical assistance and John Phillips for assistance with statistical analyses.
Disclaimer
Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Disclosure Statement
No competing financial interests exist.