
Abstract
High-pressure processing (HPP) has been proposed as an alternative quarantine method against the Mexican fruit fly Anastrepha ludens Loew (Diptera: Tephritidae), which is one of the most important pests infesting mangoes, citrus, and other fruits in Mexico and other Latin-American countries. However, processing conditions used to destroy eggs and larvae also affect the shelf life of fruits. The objective of this study was to assess the biological viability of A. ludens eggs treated with HPP, establishing whether undestroyed eggs were able to develop larvae, pupae, and adults capable of reproduction. Eggs of 1, 2, 3, and 4 days old were pressurized at 50, 75, or 100 MPa for 0, 5, 10, or 20 min. The hatching ability of pressurized eggs; survivorship of third instars, pupae, and adults emerged; and their capability to produce viable eggs were examined. The results showed that the hatching capacity of eggs and the larval development were affected negatively by the treatment duration and level of pressure. Treatments with 100 MPa for 20 min inhibited the hatching capacity of eggs of 2, 3, or 4 days old by 100%, but the inhibition was of 99.8% for 1-day-old eggs. Most of the eggs that survived the treatments were able to produce adults that can reproduce. The percent of hatching of eggs of A. ludens oviposited for adults obtained from pressurized 1-day-old eggs treated with 100 MPa for 20 min was 64.81%. Thus, more efforts must be addressed to destroy eggs and larvae during HPP because surviving organisms can reach adult stage and reproduce.
Introduction
M
Many countries require that fruit exporters perform postharvest sanitation treatments to avoid infestation of their cultivars with nonnative fruit fly species (Gazit et al., 2004). Approved treatments include fumigation with toxic compounds, irradiation, and physical treatments using heat or cold (Armstrong and Mangan, 2007). All these treatments have demonstrated to be efficient in killing eggs and larvae developed inside the living host, with a minimal negative effect on the fruit, reasonable cost, and minimal obstacle for the marketing system (Mangan and Hallman, 1998).
Emerging technologies that have been considered useful to inactivate food microorganisms have also been studied for their potential as quarantine treatments: pulsed electric field (Hallman and Zhang, 1997), radio frequency (Wang et al., 2007), water-assisted radio frequency heat treatment (Monzon et al., 2007; Tiwari et al., 2008), microwave-vapor heat treatment (Varith et al., 2007), and high-pressure processing (HPP) (Butz and Tauscher, 1995; Neven et al., 2007; Candelario-Rodríguez et al., 2009).
Although HPP has been reported as an alternative quarantine process for different insects infecting fruits, including the Mediterranean fruit fly (Ceratitis capitata Wiedemann) (Butz and Tauscher, 1995), the Western cherry fruit fly (Rhagoletis indifferens Curran), the codling moth (Cydia pomonella L.) (Neven et al., 2007), and the Mexican fruit fly (Anastrepha ludens Loew) (Candelario-Rodríguez et al., 2009), the conditions required to destroy the pest affect the fruit (Neven et al., 2007). Recently, it has been reported that fruits are affected by HPP depending on their grade of ripeness. Green mangoes showed minimal external changes even at 100 MPa, but to kill Mexican fruit fly, which infects several fruits, including citrus, avocadoes, and mangoes, from extreme southern Texas to Costa Rica (Hallman et al., 2005), pressure levels above 150 MPa are required (Candelario-Rodríguez et al., 2009).
The efficiency of HPP was improved by combining HPP with cold (Velazquez et al., 2009a) or heat (Velazquez et al., 2009b). At an initial temperature of 50°C, eggs were more resistant than larvae to this combined treatment (Velazquez et al., 2009b). Quarantine treatments might be viable if surviving organisms lose their capability of growing up or reproducing. However, there is scarce information regarding the effect of HPP and heat on the viability of surviving eggs to produce adults that can reproduce. The objective of this work was to determine the effect of HPP at initial 50°C on the biological viability of A. ludens eggs, establishing whether undestroyed eggs were able to develop larvae, pupae, and adults capable of reproduction.
Materials and Methods
The experiments were conducted in the facility of Moscafrut at Metapa de Domínguez, Chiapas (Mexico), where A. ludens are produced routinely. The eggs used in this study were supplied by this facility and reared as reported elsewhere (Candelario-Rodríguez et al., 2009; Velazquez et al., 2009a).
High hydrostatic pressure system
Pressure treatments were performed using Cold Isostatic Press Model CIP42260 (Avure Autoclave Systems, Columbus, OH), with a high-pressure chamber having 101.6 mm inner diameter, 584.2 mm length, and 350 MPa operating pressure capacity. A mixture of 5:1 water:anticorrosive lubricant (Hydrolubric 120-B; EF Houghton, Valley Forge, PA) was used as the high-pressurizing fluid, and the temperature of this mixture was adjusted to 50°C before high pressurizing. The isostatic press equipment had no temperature control; thus, the temperature changed according to the duration of pressurization, the pressure level, and the nature of the samples introduced.
Eggs were pressurized at 50, 75, or 100 MPa for 0, 5, 10, or 20 min using an initial temperature of 50°C. The duration was started to measure when the pressure of treatment was reached. Therefore, treatments with 0 min duration were those that reached the pressure of treatment and they were immediately released. The time required to reach every selected pressure was 17, 44, or 60 sec, respectively. The duration for release of pressure was always less than 10 sec. After pressurizing eggs for 0, 5, 10, or 20 min, the final temperatures were 50°C, 48°C, 45°C, and 40°C, respectively.
The temperature and pressure levels of the treatments were selected on the basis of previous studies that have shown that 50°C is needed (Velazquez et al., 2009b) and mangoes are not resistant to HPP above 100 MPa (Candelario-Rodríguez et al., 2009). HPP needs short process time to be profitable (Velazquez et al., 2005a, 2005b). Therefore, short duration (0–20 min) was selected to make the process feasible and to minimize fruit damage.
A treatment at 50 MPa and 0°C was also performed for comparative reasons. An aliquot of egg/water solution containing 896 eggs/mL was taken to fill up completely one Eppendorf tube of 2 mL capacity. The tube was introduced into the Cold Isostatic Press and pressurized at 50 MPa for 10 min with an initial temperature in the pressurizing chamber of 0°C.
High hydrostatic pressure treatment on eggs
Eggs 1, 2, 3, or 4 days old were processed independently in two different occasions. An aliquot of egg/water solution containing 896 eggs/mL was taken from a 20 L plastic container to fill up completely two Eppendorf tubes of 2 mL capacity each. They were closed immediately to avoid the presence of air bubbles. Both tubes were introduced into the Cold Isostatic Press and pressurized in the range of 50–100 MPa for 0–20 min using an initial temperature of 50°C as described above. The concentration used (896 eggs/mL) was the usual concentration in the production of the plant and it is in the range used in the other studies (Shellie and Mangan, 2000; Neven et al., 2007).
Determination of egg hatching
After pressurizing treatments, the eggs from one of the Eppendorf tubes were extracted using a brush and then were arranged in three rows of 100 eggs in Petri dishes using a stereoscope and incubated at 26 ± 1°C for 7 days. This duration is enough to allow all viable eggs to hatch. Three replicates of this bioassay were performed.
Determination of capability of surviving eggs to form larvae
The pressurized eggs of the second Eppendorf tube were transferred using a micropipette into a humid chamber system, which consisted of a sponge impregnated with chlorinated water covered with a piece of cloth inside a Petri dish. Eggs were deposited on the surface of the cloth and incubated at 26 ± 1°C for 4 days. Eggs were recovered gently and dispersed in 300 g of diet and placed in a 1 L rectangular plastic container. Diet for larvae is prepared daily in the plant with the following formulation: water (58.36%), corncob powder (19%), inactive dry yeast (7%), corn flour (5.30%), cane sugar (9.20%), guar gum (0.10%), citric acid (0.44%), sodium benzoate (0.40%), and methylparaben (0.20%).
The containers were covered with a cloth mesh and the eggs were held at different temperatures to allow optimal larvae development: at 27°C for the first instar (1–3 days old), 26°C for the second instar (4–6 days old), and 25°C for the third instar (7–9 days old). Containers were maintained under complete darkness and 75 ± 5% relative humidity (RH). After the ninth day, the living larvae were counted and surviving percent was calculated as
Determination of surviving eggs' capability to pupate
The living larvae were transferred to Petri dishes containing vermiculite to induce pupation. They were held at 20°C and 80% RH in darkness for 48 h. After the holding period, the number of pupae formed was recorded. Pupating percent was calculated as
Determination of surviving eggs' capability to produce adults
After 48 h, the formed pupae were transferred to a chamber of maturation and held for 12 days in darkness at 26 ± 1°C and 90% RH. Pupae were separated of the vermiculite and transferred to recipients designed for inducing the emergence of adults (Candelario-Rodríguez et al., 2009). These recipients were introduced into rectangular cages of 30 cm long, 10 cm wide, and 10 cm deep, containing inside special diet for larvae (protein hydrolysate and sugar in proportion of 1:3) and water. Adults began to emerge after 5 days, and the number of adults was recorded. The percent of adults emerged was calculated as
Determination of reproductive capacity of emerged adults
Adults emerged from pressurized eggs were allowed to reproduce. If only one adult was obtained from the pressurizing treatment or if only adults of the same sex were obtained, fertile adults of the same age obtained from nonpressurizing treatments were introduced into the cages in a proportion of 1:3 (emerged from pressurizing treatment:no treated adults).
The ovipositor panels were installed after 10 days from the emergence of the adults, because it is the time when adults reach the sexual maturity. Every 2 days, the eggs were collected and the hatching was determined as described above. The ability of eggs to hatch was determined five times for each treatment. All rearing procedures and obtaining data from egg hatching to adult oviposition were adapted from the Manual of Mass Rearing, Planta Moscafrut (2009).
Microscopic photography of eggs
Nonhatching eggs were photographed using a digital camera (DXM1200F; Nikon Instruments Inc., Melville, NY) attached to a dissecting microscope (SM21500; Nikon).
Statistical analysis
Statistical analysis was performed using Statgraphics 5.0 (Software Publishing Corporation–Bitstream, Inc., Cambridge, MA). Least significant difference's multiple range tests were used to determine significant differences (p < 0.05) among treatments.
Results
Effect of high hydrostatic pressure on eggs
The effect of the pressure level and duration of treatment at initial 50°C on the hatching ability of A. ludens eggs is shown in Table 1. The hatching capacity of eggs was affected by the treatment duration and pressure level. The effect of duration of treatment (0–20 min) was more evident at 50 MPa, and the effect of pressure level (50–100 MPa) was more evident at 0 min. The hatching ability of eggs from all ages was less affected by pressurizing them at 50 MPa for 0 min, but 3-day-old eggs were less sensitive to this treatment. This behavior was also observed in 3-day-old eggs at 75 MPa for 0 min.
Mean values and standard deviations of two studies with three replicates. Values are percentages of eggs that did not hatch in each treatment.
Different letters in the same row indicate significant differences (p < 0.05) between treatment durations for each level of pressurizing.
The hatching rate was highly decreased when pressure levels of 50–100 MPa were applied for 10–20 min. Most of the eggs pressurized at 100 MPa for 10 or 20 min lost almost completely their ability of hatching. However, some 1-day-old eggs were able to hatch, forming larvae at 100 MPa even after 20 min.
Figure 1 shows the effect of the HPP on the inner structure of A. ludens eggs that were not able to hatch compared with nontreated eggs. The eggs pressurized at 50 MPa and 50°C for 10 min (Fig. 1b) showed an inner structure with evident flocculation and fragmentation of the proteinaceous structure with a separation of the solid phase respect to the liquid phase. Similar behavior was observed in the eggs pressurized at 50 MPa but at 0°C (Fig. 1c).
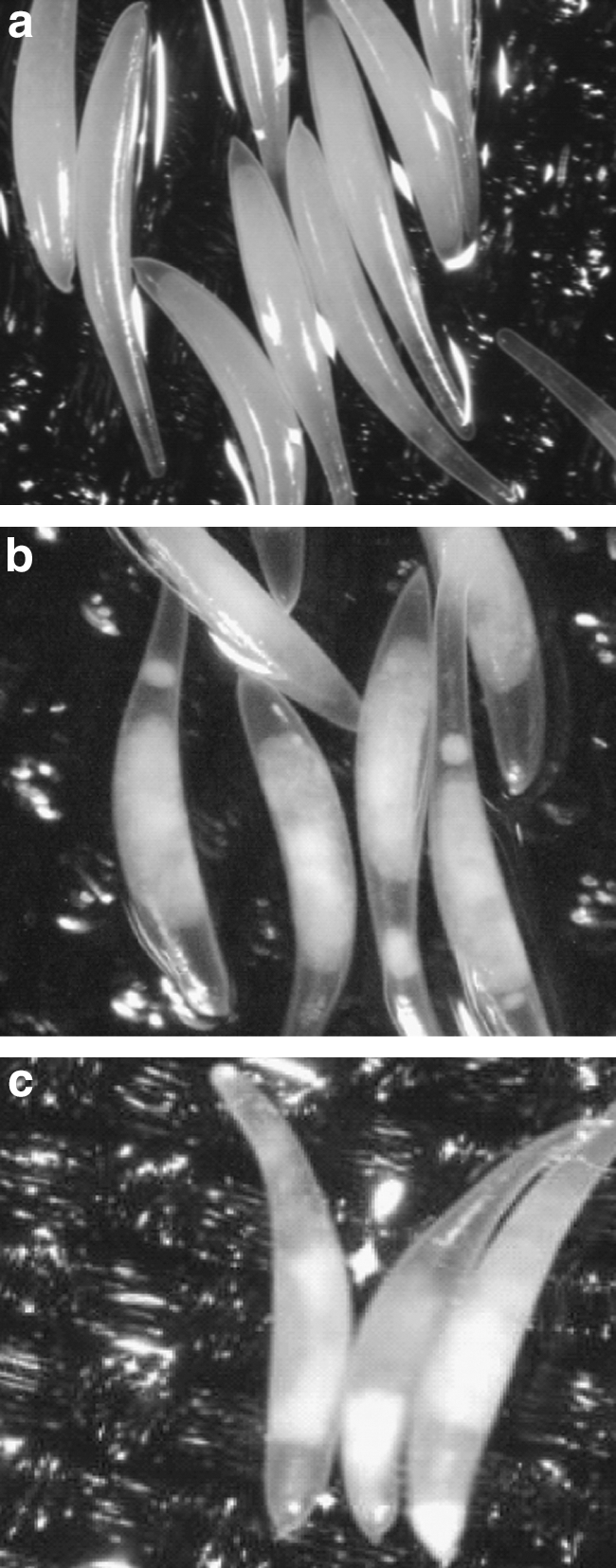
Microscopic photography of 1-day-old eggs of Anastrepha ludens. (
Development of pressurized eggs to form larvae, pupae, and adults
The effect of treatments to inhibit the ability of eggs to form larvae and the viability of such larvae to grow up to reach the third instars after 9 days of incubation was followed, and the results are shown in Table 2. The rate of larvae formed from pressurized eggs and survived until third instars was affected by both treatment duration and pressure level.
Mean values and standard deviations of two studies with three replicates.
Different letters in the same row indicate significant differences (p < 0.05) between treatment durations for each level of pressurizing.
Eggs pressurized at 50 MPa for 0 min were able to produce a higher number of larvae, but the number of larvae formed from eggs pressurized at 50 MPa decreased by increasing treatment duration. The same behavior was observed at 75 MPa. The ability to form larvae decreased to the minimum by pressurizing eggs at 100 MPa even for 0 min. Note that even at 100 MPa for 20 min, some pressurized eggs were able to hatch and form larvae with appropriate physiology to grow up until reach the third instars.
The effect of HPP on the ability to pupate of larvae from surviving eggs was determined and results are shown in Table 3. Eggs pressurized at 50 MPa for 0 min were able to produce a few number of larvae with the ability to reach the stage of pupa, but the number of pupae formed from eggs pressurized at 50 MPa decreased by increasing the treatment duration. The same behavior was observed at 75 MPa. The ability to form pupa decreased to the minimum by pressurizing eggs at 100 MPa even for 0 min, but even at 100 MPa for 20 min, some pressurized eggs were able to form larvae with appropriate physiology to grow up until reach the pupa stage.
Mean values and standard deviations of two studies with three replicates.
Different letters in the same row indicate significant differences (p < 0.05) between treatment durations for each level of pressurizing.
HPP affected the ability of eggs to hatch and reach the adult stage, but some specimens were able to survive the treatments and preserve their ability to form adults (Table 4). Eggs pressurized at 50 MPa for 0 min were able to produce a few numbers of adults, but the number of adults formed decreased by increasing the treatment duration. The same behavior was observed at 75 MPa. The ability to form adults decreased to the minimum by pressurizing eggs at 100 MPa even for 0 min. Some pressurized eggs were still able to form larvae with appropriate physiology to grow up until reach the adult stage at 100 MPa for 20 min.
Mean values and standard deviations of two studies with three replicates.
Different letters in the same row indicate significant differences (p < 0.05) between treatment durations for each level of pressurizing.
Reproductive ability of adults obtained from pressurized eggs
The adults obtained from pressurized eggs were matched with individuals of their opposite sex to determine whether they preserved their reproductive ability and whether the new eggs were able to hatch (Table 5). Although some individuals from specific treatments were not able to reproduce, most of the flies obtained from pressurized eggs were able to reproduce and their eggs were able to hatch. Adult fly specimens that did not reproduce were 2-day-old eggs pressurized at 50 MPa for 10 min and at 100 MPa for 5 min; 3-day-old eggs pressurized at 75 MPa for 20 min; 4-day-old eggs pressurized at 100 MPa for 20 min. Even adults obtained from eggs pressurized at 100 MPa for 20 min, the maximum processing condition for this study, were able to reproduce and their eggs preserved their ability to hatch.
Mean values and standard deviations of two studies with five replicates.
Discussion
Effect on eggs hatching
In this study, some treatments that should be more lethal (high pressure and long duration) showed less lethality than other treatments that are less stressing. This difference might be caused by a decrease in the temperature of the vessel during the process (from 0°C to 10°C depending on time), since HPP is instantaneous and uniform in a closed system (Téllez-Luis et al., 2001; Torres and Velazquez, 2005; Velazquez et al., 2005a, 2005b). On this regard, it has been reported that the resistance to heat of fruit fly eggs from different species, including A. ludens, decreases strongly above 40°C. At 44°C, 50–80 min was required to obtain 100% mortality depending on the species, but at 50°C only 0.5–3 min was required (Armstrong et al., 2009). Despite the variation in temperature, some eggs showed higher resistance to the HPP than other those affected by some thermal treatment. This resistance could be associated with physiological properties of individuals.
Eggs pressurized at 50 MPa for 10 min showed a different inner structure from control nonpressurized eggs (Fig. 1). Note that microphotography was obtained from the eggs that did not hatch because eggs that hatched formed larvae and could not be observed. Eggs are composed mainly for water and proteins. Protein denaturation/aggregation induced by high pressure depends on the pressure level, pressure holding time, and temperature (Borderías et al., 1997; Fernández-Martín et al., 1998; Uresti et al., 2004).
The adverse effect of HPP on egg hatching has been associated with a negative effect on the structure and permeability of external and internal membranes, a negative effect on proteins (denaturating/aggregation), or a combination of both factors (Velazquez et al., 2009a). In the present study, it was observed that eggs pressurized at 50 MPa and 50°C for 10 min showed the presence of protein flocculation and separation of the proteinaceous structure, as compared with control nonpressurized eggs (Fig. 1). Although both pressure and heat treatment are capable of inducing flocculation of egg proteins, eggs pressurized at the same conditions but at 0°C showed a similar behavior, having their protein flocculated and the proteinaceous mass disaggregated (Fig. 1c). Protein denaturation/aggregation induces a different damage that affects the viability of the organism (Farr, 1990; Pothakamury et al., 1995; Téllez-Luis et al., 2001).
The resistance of some specimens to HPP might be explained as the response of some specimens having tolerance to cold or heat shock injury and tolerance to long-term heating and chilling injury (Denlinger and Lee, 1998; Denlinger and Yocum, 1998). Resistance to HPP of some organisms might be associated with the presence or production of baroprotective solutes surrounding the proteins. Sugars and polyols at 8%–12% have a stabilizing effect on proteins against high pressure (Ashie et al., 1999; Uresti et al., 2005).
Surviving larvae obtained from pressurized eggs
The number of surviving larvae that reached the third instars (Table 2) was lower than the number of eggs hatched (Table 1). This difference might be associated with physicochemical changes caused by the pressurizing treatment on proteins involved in the DNA replication and transcription, inducing biochemical and genetic changes (Farr, 1990; Pothakamury et al., 1995; Téllez-Luis et al., 2001). Such changes might be responsible for inducing the formation of larvae with physiological defects that inhibit growth to reach third instars. However, it is important to consider natural mortality (lower than 10%) and the difference in the methodology, because hatching was determined as a probability by taking out a sample, whereas surviving larvae were obtained from the totality of the sample.
Formation of pupae from surviving larvae
Although some larvae were not able to pupate, most of them did (Table 3), indicating that most of the eggs that survived to HPP preserved their physiological properties and biological functions associated with growing and evolution. Recently, it was shown that A. ludens larvae were able to resist and pupate even after being pressurized at 150 MPa for 20 min at 25°C (Candelario-Rodríguez et al., 2009), 150 MPa for 10 min at 0°C (Velazquez et al., 2009a), and at 75 MPa for 0 min at 50°C (Velazquez et al., 2009b).
Formation of adults from pupae
Most of the pupae emerged into adult flies, confirming that most of the eggs that survived HPP preserved their physiological properties and biological functions associated with growing and evolution. Butz and Tauscher (1995) found that eggs of Mediterranean fruit fly that survived treatment of 10–20 min at 100 MPa and 25°C were able to hatch, form larvae and pupate, and subsequently develop adults.
Viability of eggs produced by adults obtained from pressurized eggs
Most of the adults showed the ability to reproduce and produce viable eggs (Table 5). These results indicate that efforts to optimize HPP as a quarantine method for A. ludens must be focused to destroy both eggs and larvae during treatment because most of the surviving organisms are able to form adults that preserve their reproductive capacity.
Practical considerations
HPP has been reported as an alternative quarantine process for the Mediterranean fruit fly (Ceratitis capitata Wiedemann) (Butz and Tauscher, 1995), the Western cherry fruit fly (Rhagoletis indifferens Curran), the codling moth (Cydia pomonella L.) (Neven et al., 2007), and the Mexican fruit fly (Candelario-Rodríguez et al., 2009). However, pressure conditions required to destroy the targeted pest affect the host fruit (Neven et al., 2007; Candelario-Rodríguez et al., 2009).
Recently, it has been reported that combining HPP with low or high temperature is feasible to reduce the level of pressure required to destroy eggs or larvae of A. ludens (Velazquez et al., 2009a; 2009b). These results allow considering HPP as an alternative quarantine method, but more studies are essential to obtain conditions under 100 MPa, which is the pressure level that unripe mangoes seem to tolerate well (Candelario-Rodríguez et al., 2009).
Conclusions
HPP at initial 50°C was able to destroy most of the A. ludens eggs at 50–100 MPa with 10–20 min of treatment. However, some eggs were able to survive the process and preserve the ability to hatch, form larvae, pupate, and develop into adults with reproductive capacity. Egg mortality was associated with the induction of denaturation/aggregation of the cytoplasm protein. Surviving of some eggs seems to be associated with the presence of special defensive mechanism of specimens perhaps associated with genetic heritage. Thus, efforts must be addressed to destroy all eggs and larvae during HPP because most of the surviving organisms can reach adult stage and reproduce.
Footnotes
Acknowledgments
The authors are grateful to Programa de Mejoramiento del Profesorado for financial support to this work (Red Aprovechamiento de Recursos Agropecuarios), and to CONACYT for the scholarship granted to Hugo Ernesto Candelario Rodrìguez and Juan Francisco Castañón Rodrìguez. We are also grateful to José Manuel Gutiérrez Ruelas, Director de Moscas de la Fruta, DGSV-SENASICA SAGARPA, for logistic support, and Refugio Hernández and Emilio Hernández for technical and scientific advice.
Disclosure Statement
No competing financial interests exist.