
Abstract
Yersinia enterocolitica is a foodborne pathogen well known for its ability to grow at low temperatures. Recent studies with another psychrotrophic foodborne pathogen, Listeria monocytogenes, revealed that temperature of growth had pronounced impact on survival following repeated freezing and thawing (cryotolerance). Listerial cryotolerance was significantly more pronounced when bacteria were grown at 37°C than following growth at either 4°C or 25°C. However, it is not known whether such impact of growth temperature is a general adaptation shared with other foodborne pathogens. In this study, we investigated the impact of growth temperature (4°C, 25°C, and 37°C) on cryotolerance of Y. enterocolitica. In strong contrast to findings previously obtained with Listeria spp., cryotolerance of Y. enterocolitica was impaired following growth in liquid media at 37°C, with cell concentration dropping to undetectable levels (<101 colony forming unit/mL) following as few as six freeze–thaw cycles. On the other hand, when the bacteria were grown at 4°C, cryotolerance was significantly higher (p < 0.05), and substantial survival was maintained even after 18 cycles (2–5 log reduction, depending on strain). Enhanced cryotolerance was also observed with cultures grown at 25°C. Bacteria grown at 37°C on agar were significantly more cryotolerant than following growth at 37°C in liquid media (p < 0.05). The data suggest species-specific impact of growth temperature and liquid versus agar medium on cryotolerance of cold-tolerant bacteria.
Introduction
P
Yersinia enterocolitica is a Gram-negative bacterium associated with significant human disease burden in the United States and other nations, and transmitted via contaminated food or water (Bottone, 1997; Fredriksson-Ahomaa et al., 2006). Swine is the major reservoir for strains implicated in human yersiniosis, and Y. enterocolitica can be isolated from raw pork products (Bottone, 1997; Fredriksson-Ahomaa et al., 2006; Wesley et al., 2008). Y. enterocolitica has a wide growth temperature range (−5°C to 42°C) and are well known for their ability to grow at low temperatures (Brocklehurst and Lund, 1990; Robins-Brown, 1991). Extensive data indicate that temperature has pronounced and complex impact on expression of virulence determinants and on the pathogen's interactions with host cells (Bottone, 1997; Revell and Miller, 2000). In contrast, the impact of growth temperature on the response of Y. enterocolitica to environmental stresses remains poorly understood.
Raw meat products are frequently frozen for preservation, and several studies have addressed Y. enterocolitica's survival following freezing, with variable results (Bhaduri, 2005, 2006; Hanna et al., 1977). Y. enterocolitica can survive freezing remarkably well, with no loss in survival or detectable cell injury after 12 weeks in frozen ground pork (Bhaduri, 2005). However, limited information is available on the impact of different growth temperature and of repeated freezing and thawing on survival of Y. enterocolitica. We wished to determine whether growth of Y. enterocolitica at a temperature associated with mammalian infection (e.g., 37°C) may be also required for cryotolerance, as was observed with L. monocytogenes (Azizoglu et al., 2009). Further, it was important to determine whether cryotolerance may be impacted by planktonic versus surface-associated growth of the bacteria before freezing. In this study we investigated freeze–thaw tolerance of Y. enterocolitica grown at 4°C, 25°C, and 37°C, following repeated cycles of freezing (−20°C) and thawing.
Materials and Methods
Bacterial strains and culture conditions
Y. enterocolitica isolates used in this study are listed in Table 1 and included two strains of human origin, YE 9610 (biovar 1, serogroup O:8) and SK 3181, and three from pork tongue. Bacteria were grown on trypticase soy agar with 5% sheep blood (Remel, Lenexa, KS) at 37°C for 36 h and liquid cultures were started by transferring a single colony into 5 mL of trypticase soy broth (TSB; BBL, Cockeysville, MD) supplemented with 0.7% Yeast Extract (YE; Becton, Dickinson & Co., Sparks, MD) (TSBYE) and incubating at 37°C overnight. Thirty microliters of this culture was added to 30 mL of TSBYE and incubated at 4°C, 25°C, or 37°C for 3 weeks, 48 h, or 36 h, respectively, to stationary phase (OD600 ∼1.0; OD600 measured with a spectrophotometer [SmartSpec 3000; Bio-Rad, Hercules, CA]). Colony forming units (CFU)/mL were determined by plating in duplicate on TSBYE containing 1.5% agar (Becton, Dickinson & Co.) after serial dilution in TSBYE, and incubation at 37°C for 36 h. To determine the impact of growth on solid medium on cryotolerance, the bacteria were grown on TSBYE containing 1.5% agar at 37°C for 48 h, and the colonies were swabbed by a sterile cotton swab (Fisher Scientific, Houston, TX), and suspended in 10 mL of TSBYE, with OD600 adjusted to ∼1.0, to ensure comparable initial cell counts between cultures grown in broth and on agar.
Freezing and thawing treatments and assessment of cryotolerance
Bacterial suspensions from cultures grown in broth or on agar as described above were transferred (1.5 mL) into sterile cryovials (Nalgene, Rochester, NY) and frozen at−20°C. Thawing was performed for 10 min in a room temperature water bath. Freezing and thawing cycles were repeated every 24 h for 18 cycles. Every three cycles, enumerations of CFU/mL were done in duplicate, as described above. Freezing rates were determined with thermocouples, as described previously (Azizoglu et al., 2009).
Statistical analysis
All treatment combinations were replicated twice. Log reduction of each sample was determined following 18 freeze–thaw cycles. The log reduction data of each strain–temperature combination were analyzed by performing analyses of variance. The significance was determined at unadjusted level of α = 0.05. All statistical analyses were performed using SAS c.9.1 (Cary, NC).
Results
Enhanced freeze–thaw tolerance in Y. enterocolitica grown planktonically at 4°C or 25°C, in comparison to 37°C
Y. enterocolitica grown at 37°C in liquid underwent pronounced reductions in CFU following repeated freezing and thawing, with CFU/mL after six freeze–thaw cycles ≤101. Even a single freeze–thaw cycle of bacteria grown in broth at 37°C resulted in 2–3 log reductions (Fig. 1A–C). On the other hand, when the bacteria were grown in liquid at 4°C, cryotolerance was noticeably higher for all tested isolates (p < 0.05), with <1 log reduction following six freeze–thaw cycles (Fig. 1A–C). Differences in survival were noted among different isolates following additional freeze–thaw cycles. After 18 freeze–thaw cycles YE 9610 and SK 3181 had ∼2.5–4 log reduction, whereas the pork isolate YE 14A had <1 log reduction (p < 0.05) (Fig. 1A–C). Similarly, low reductions (<1 log) were observed with the other two pork isolates, YE 7A and YE 10A, following 18 freeze–thaw cycles (data not shown).
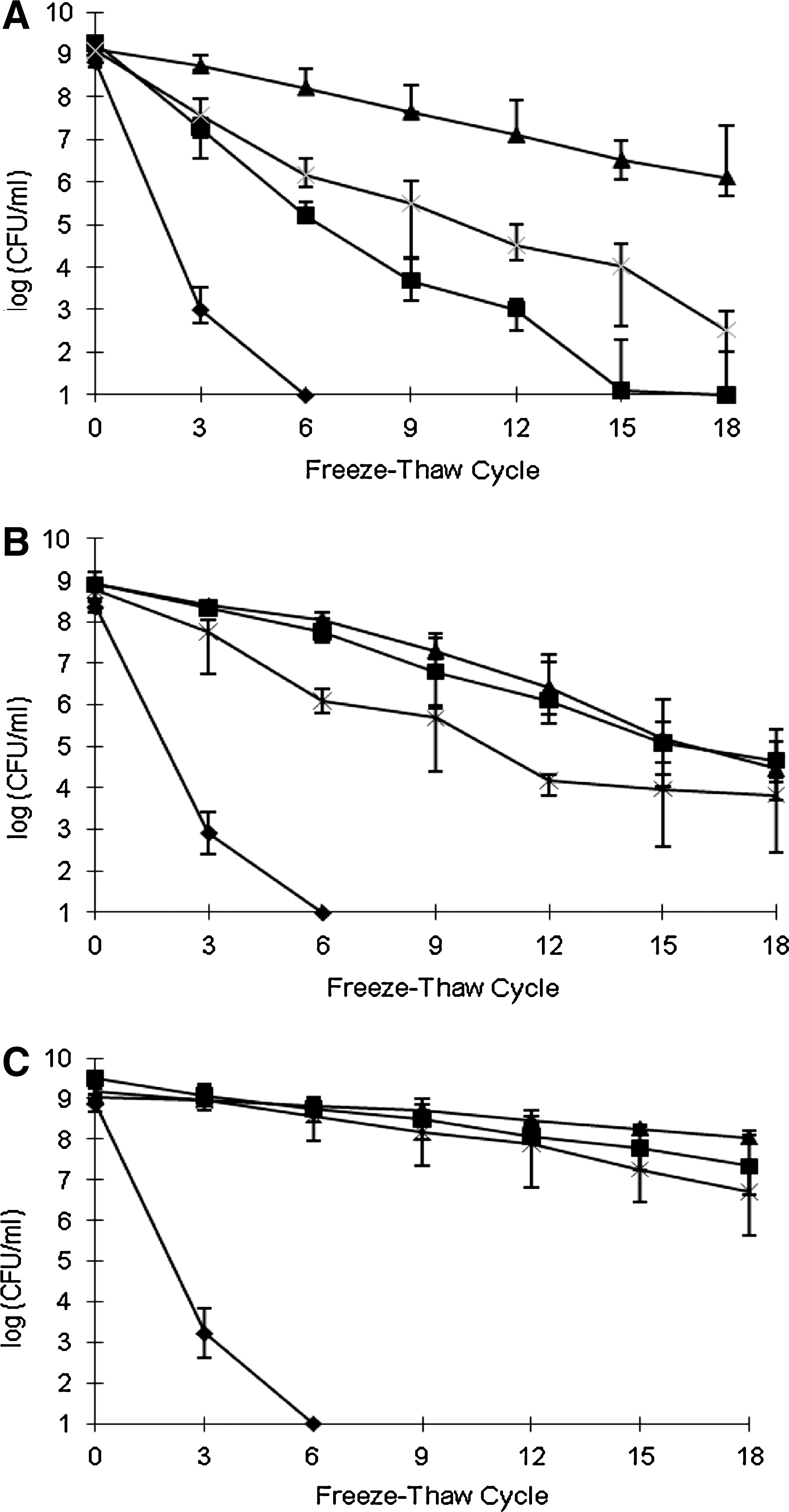
Impact of growth temperature and agar versus liquid medium on freeze–thaw tolerance of Yersinia enterocolitica. Y. enterocolitica isolates were YE 9610 (
Growth of the bacteria at 25°C also conferred increased cryotolerance in comparison to 37°C-grown cultures. In the case of YE 9610, freeze–thaw tolerance of 4°C-grown cells was still significantly higher than that of cells grown at 25°C (p < 0.05) (Fig. 1A). On the other hand, for SK 3181 and the pork isolates YE 7A, YE 10A, and YE 14A, the protective effect of growth at 25°C on cryotolerance was similar to that conferred by growth at 4°C (p > 0.05) (Fig. 1B, C, and data not shown).
Protective impact on cryotolerance of growth on solid media at 37°C
In contrast to the rapid reductions in CFU following repeated freezing and thawing of cells grown at 37°C in liquid, when the cultures were grown at 37°C on agar their freeze–thaw tolerance was markedly enhanced (p < 0.05) (Fig. 1A–C). Protective impact of growth on agar was noted with all three tested strains (YE 9610, SK 3181, and YE 14A), although strain-specific differences were noted. Cryotolerance of YE 9610 grown at 37°C on agar was significantly higher than of cells grown in liquid at 37°C, and was even higher than the freeze–thaw tolerance of cells grown in liquid at 25°C. Nonetheless, YE 9610 grown at 37°C on agar was still more sensitive to repeated freezing and thawing than following growth at 4°C in liquid (Fig. 1A). Superior cryotolerance of 4°C-grown cells was also observed with SK 3181 (p < 0.05) (Fig. 1B). YE 14A, which exhibited the highest cryotolerance following 4°C growth, also had the highest enhancement in cryotolerance of the cells following growth at 37°C on agar, in comparison with cells grown planktonically at 37°C (p < 0.05) (Fig. 1B). In fact, cryotolerance of YE 14A grown on agar at 37°C was not significantly different from that of cells grown in liquid at either 4°C or 25°C (p > 0.05) (Fig. 1C).
Discussion
In this study, cryotolerance of all tested isolates of the psychrotrophic microorganism Y. enterocolitica was significantly lower with cells grown planktonically at 37°C than cells grown at 4°C or 25°C. The protective impact of low-temperature growth was strain specific, being more pronounced for the three meat isolates than for the two strains of clinical origin. Growth at 37°C on agar was associated with significant enhancements in cryotolerance in comparison to cells grown planktonically at 37°C.
The overall impact of growth temperature in planktonically grown cells was in marked contrast to recent findings with L. monocytogenes, for which cryotolerance was significantly higher when cells were grown in liquid media at 37°C than at 4°C and 25°C (Azizoglu et al., 2009). Instead, our findings with Y. enterocolitica are reminiscent of those obtained with Exiguobacterium spp. isolated from permafrost and other environments. Exiguobacterium spp. had markedly higher cryotolerance following planktonic growth at 4°C than at higher temperature (25°C), and growth at 25°C on agar had a protective impact on cryotolerance (Vishnivetskaya et al., 2007), similar to that observed for 37°C agar-grown cells of Y. enterocolitica. Growth at 4°C (vs. 25°C) was also found to enhance survival of other bacteria from Siberian permafrost following freezing for prolonged periods (Ponder et al., 2005).
Freezing and thawing involves a number of different stresses, including cold, osmotic, and oxidative stress (Mazur, 1970; Gao and Critser, 2000). In Campylobacter, coping with oxidative stress was shown to be required for freeze–thaw tolerance (Stead and Park, 2000; Garénaux et al., 2008). Thus, the observed opposite impact of growth temperature on the cryotolerance of L. monocytogenes compared to Y. enterocolitica or Exiguobacterium spp. might reflect differences in the ability of the bacteria to cope with oxidative stress following growth at different temperatures. In this context, the protective impact of surface-associated growth (observed in Y. enterolitica as well as Exiguobacterium spp., and for at least one strain–temperature combination in L. monocytogenes [Vishnivetskaya et al., 2007; Azizoglu et al., 2009]) may also reflect the increasingly recognized impact of planktonic versus surface growth on the physiology of microorganisms. In the case of Y. enterocolitica, surface antigenic changes in response to growth temperature have been noted (Kawaoka et al., 1982; Wartenberg et al., 1983), but the dependence of such changes on planktonic versus surface growth is not known. Further studies will be needed to assess the involvement of these and other mechanisms involved in low-temperature adaptations (Annamalai and Venkitanarayanan, 2005, 2009; Goverde et al., 1998; Neuhaus et al., 2003; Palonen et al., 2010) on cryotolerance of these microorganisms.
Even though the mechanisms mediating the observed impact of growth temperature on cryotolerance of Y. enterocolitica remain to be identified, such impact may be relevant to the long recognized hot–cold cycle of transmission of this psychrotrophic pathogen (Bottone, 1997). Limited cryotolerance of bacteria freshly excreted from warm-blooded animals (e.g., swine) into the environment could result in population bottlenecks; survivors would subsequently grow at low temperature, and acquire enhanced tolerance to freezing and thawing, thus promoting environmental persistence of the pathogen. Such population bottlenecks and regrowth may also contribute to the genetic variation of the organism in its environmental reservoirs.
In our study we noted significant differences in cryotolerance, and in the protective effect of surface growth, among different isolates. The reproducibility of the results suggests that the observed differences have a genetic basis, the components of which remain to be identified. Studies including isolates from diverse sources will be needed to assess the extent to which cryotolerance may differ between environmental and clinical isolates, or among isolates from different regions. Further studies are also needed to assess the possible impact of growth temperature on cryotolerance of other pathogenic Yersinia spp., including Y. pestis and the foodborne pathogen Y. pseudotuberculosis.
Footnotes
Acknowledgments
This project was partially funded by the U.S. Department of Agriculture Grant 2006-35201-17377. We thank Dr. K. Venkitanarayanan and Dr. L. Wolf for strains. We are grateful to R.M. Siletzky for laboratory support and to all other members of our laboratory for discussions, encouragement, and support in the course of this project.
Disclosure Statement
No competing financial interests exist.