
Abstract
High concentration of copper, fed as copper sulfate, is often used to increase growth rates in swine. Bacteria exposed to copper may acquire resistance, and in Enterococcus faecium and Enterococcus faecalis, a plasmid-borne transferable copper resistance (tcrB) gene that confers copper resistance has been reported. Our objectives were to determine the occurrence of tcrB in fecal enterococci from weaned piglets fed diets with a normal supplemental level (16.5 ppm) or an elevated supplemental level (125 ppm) of copper and to determine the association of tcrB with copper, erythromycin, and vancomycin resistance. A total of 323 enterococcal isolates were examined and 15 (4.6%) isolates (14 E. faecium and 1 E. faecalis) were positive for tcrB. Fifteen tcrB-positive and 15 randomly chosen tcrB-negative isolates from piglets fed the normal supplemental level of copper were tested for erm(B), tet(M), vanA, and vanB genes and susceptibilities to copper, erythromycin, tetracyclines, and vancomycin. All tcrB-positive and -negative isolates contained erm(B) and tet(M), but not vanA and vanB. The mean minimum inhibitory concentration of copper for tcrB-positive (21.1 mM) was higher (p < 0.001) compared with tcrB-negative isolates (6.1 mM). All isolates were resistant to erythromycin and tetracyclines and susceptible to vancomycin. The transferability of the tcrB gene from tcrB-positive strains to tcrB-negative strains was demonstrated by conjugation. The potential link between tcrB and antibiotic resistance genes and the propensity of enterococci to transfer tcrB to other strains raises the possibility that copper supplementation may exert selection pressure for antibiotic-resistant enterococci. This study is the first report on the occurrence of the tcrB gene in enterococci isolated from swine in the United States.
Introduction
C
Copper homeostasis in gram-positive bacteria, well documented in Enterococcus hirae, is mediated by a group of membrane-spanning proteins, called CPx-ATPases, which are encoded by four genes in an operon, called cop YZAB (Magnani and Solioz, 2005; Solioz and Stoyanov, 2003). The copA and copB encode for ATPases, which are responsible for influx and efflux of copper, respectively. The copZ encodes a copper chaperone and copY acts as copper responsive repressor (Solioz and Stoyanov, 2003). Acquired copper resistance has been reported in gram-negative bacteria, such as Escherichia coli and Pseudomonas syringae (Brown et al., 1995; Silver, 1996), and in certain gram-positive bacteria, E. hirae, Bacillus subtilis, and Lactococcus lactis (Leelawatcharamas et al., 1997; Hasman and Aarestrup, 2002; Solioz et al., 2010). A copper resistance gene, designated as transferable copper resistance gene or tcrB, that confers copper resistance has been identified in Enterococcus faecium, Enterococcus faecalis, Enterococcus gallinarum, Enterococcus casseliflavus, and Enterococcus mundtii (Hasman and Aarestrup, 2002; Hasman et al., 2006b). The tcrB gene, harbored on a plasmid (Hasman et al., 2006b), is homologous to copB of cop YZAB operon, and encodes a putative protein belonging to the CPx-type ATPase family (Hasman, 2005). Interestingly, the plasmid also carried genes erm(B) and vanA, which encode resistance to macrolides and glycopeptides, respectively (Hasman and Aarestrup, 2002; Hasman et al., 2006b), suggesting a potential linkage of copper resistance to antibiotic resistance. Because tcrB in enterococci has only been reported in Europe, we conducted a study to determine the occurrence of tcrB in fecal enterococcal isolates of piglets supplemented with elevated levels of copper and relate tcrB to phenotypic susceptibilities to copper, erythromycin, and vancomycin. Also, the transferability of tcrB from tcrB-positive to tcrB-negative enterococci strains was determined by conjugation (in vitro).
Materials and Methods
Animals, experimental design, and sampling
The use of animals and the experimental procedure followed were approved by the Kansas State University Animal Care and Use Committee. Fecal enterococci for this study were obtained from weaned piglets fed diets supplemented with or without an elevated supplemental level of copper supplied by copper sulfate. The study was designed and conducted to assess the performance benefits of supplementation of copper, zinc, or antibiotics. One hundred and fifty weaned pigs (21 days old with an average body weight of 6.0 ± 1.0 kg) were randomly allocated to one of five dietary treatments. The five dietary treatments were basal diet with 16.5 ppm of supplemental copper and 165 ppm of supplemental zinc (control), basal diet supplemented with 125 ppm of copper provided by copper sulfate (copper group), basal diet supplemented with 3000 ppm of zinc provided by zinc oxide (zinc group), basal diet supplemented with 125 ppm copper and 3000 ppm of zinc (copper and zinc group), and basal diet supplemented with neomycin sulfate and oxytetracycline (Neo/Oxy 10/10; Penfield Animal Health, Omaha, NE; antibiotic group) at 55 mg/kg of feed (1:1). In treatment groups that were supplemented with zinc (zinc and copper and zinc groups), the level of zinc supplementation was reduced from 3000 to 2000 ppm after 14 days (Smith et al., 1997). The levels of copper and zinc supplemented in the basal diet are typical supplementation rates provided in nursery diets to meet the nutrient needs of the piglets. The basal diet consisted of corn, soybean meal, and vitamins, amino acids, and trace mineral supplements, and piglets were housed in an environmentally controlled nursery facility. Each dietary treatment group had a total of 30 piglets assigned to 6 pens with 5 piglets per pen. Each pen contained a four-hole, dry self-feeder and a nipple water to provide ad libitum access to feed and water. Pens had wire-mesh floor and that allowed for 0.3 m2 per piglet. Piglets were fed the treatment diets for 5 weeks. During each week of the study, fecal samples were obtained from three randomly selected piglets per pen and the samples were transported on ice immediately to the laboratory.
Isolation and identification of enterococci
All culture media used in the study were from BD (Becton and Dickson, Sparks, MD). Fecal samples were diluted (∼1 g in 10 mL) in phosphate-buffered saline and 50 μL of the suspension was spread-plated onto M-Enterococcus agar. After 24 h of incubation at 37°C, two colonies (pin-point red, and pink or metallic pink) were picked, streaked on blood agar plates, and incubated overnight at 37°C. For the genus confirmation, isolates were inoculated into 100 μL Enterococcosel broth in a 96-well microtiter plate (Becton and Dickinson, Franklin Lakes, NJ), incubated at 37°C for 4 h, and tested for esculin hydrolysis, followed by testing with the Analytical Profile Index (API) 20 Strep kit (bioMérieux®, Durham, NC). Isolates confirmed as enterococci were stored in protect beads (Cryo-Vac®, Round Rock, TX) at −80°C until further use.
Polymerase chain reaction for detection of the tcrB gene
The tcrB gene in enterococcal isolates was detected by the procedure described by Hasman et al. (2006b). For DNA extraction, each isolate from the protect bead was streaked on a blood agar plate and a single colony was suspended in nuclease free water with Chelex® 100 Resin (Bio-Rad Laboratories, Hercules, CA) and boiled for 10 min. The primers (Table 1) for the polymerase chain reaction (PCR) were supplied by Integrated DNA Technologies (IDT, Coralville, IA). One of the two tcrB-positive E. faecium strains obtained from Denmark (7430275-4 or 7430162-6) served as a positive control.
Speciation of enterococci
Species identification of tcrB-positive and an equal number of tcrB-negative enterococcal isolates, randomly chosen from the control group, was performed by a multiplex PCR that identifies E. faecium, E. faecalis, E. gallinarum, and E. casseliflavus (Jackson et al., 2004a). Additionally, superoxide dismutase (sodA) gene sequence analysis (Poyart et al., 2000) was used for confirmation of species. The DNA of the isolates was extracted as described above. Master mixes, primers (Table 1), and running conditions for the multiplex PCR were as described by Jackson et al. (2004a). The ATCC strains of E. faecium (ATCC 19434), E. faecalis (ATCC 19433), E. gallinarum (ATCC 49579), and E. casseliflavus (ATCC 25788) served as positive controls. The primers (Table 1) and PCR conditions for sodA sequence analysis were as described by Poyart et al. (2000). The primers used were supplied by Invitrogen Life Technologies (Carlsbad, CA).
DNA sequencing
PCR products (tcrB and sodA) were purified by QIAquick® Gel Extraction Kit (QIAgen Science, Valencia, CA). The eluted DNA samples were submitted for sequencing to Genomics Core, Institute for Integrative Genome Biology, University of California at Riverside. The sequences were analyzed by BLAST search in the NCBI GenBank database.
Detection of erm(B), tet(M), vanA, and vanB genes
The primers (Table 1) and PCR conditions for detection of erm(B) and tet(M) genes were as per Jacob et al. (2008). E. faecalis MMH 594 and E. coli harboring plasmid pFD 310 (Aminov et al., 2001) served as positive controls for erm(B) and tet(M), respectively. The primers (Table 1) and PCR conditions used for detecting vanA and vanB genes were based on Kariyama et al. (2000). The E. faecium (ATCC 51559) and E. faecalis V583 were used as positive controls for vanA and vanB, respectively.
Copper susceptibility determinations
Copper susceptibilities of enterococcal isolates were determined by agar dilution method (Hasman et al., 2006b). The tcrB-positive isolates, including the 2 E. faecium strains from Denmark (7430162-6 and 7430275-4; provided by Henrik Hasman, National Food Institute, Technical University of Denmark) and 15 tcrB-negative strains were included. Mueller–Hinton agar plates containing 0, 2, 4, 8, 12, 16, 20, 24, 28, 32, 36, or 40 mM of copper sulfate (Fischer Scientific, Fair Lawn, NJ), adjusted to pH 7.0, were used for copper susceptibility determinations. The plates, in duplicates, were spot inoculated with 20 μL of bacterial growth that was adjusted to McFarland turbidity standard no. 0.5. Plates were incubated for 48 h at 37°C to determine growth or no growth. The susceptibility determination was repeated with different inocula preparations.
Antibiotic susceptibility determinations
Minimum inhibitory concentrations (MICs) of antibiotics were determined by micro-broth dilution method (CLSI, 2002). Antibiotics tested were chlortetracycline, erythromycin, oxytetracycline, tetracycline, and vancomycin (Sigma-Aldrich, St. Louis, MO). Stock solutions of antibiotics were prepared in sterile distilled water to obtain a concentration of 1000 μg/mL based on potency of antibiotics. Antibiotics were tested at concentrations of 100, 50, 25, 12.5, 6.25, 3.125, 1.56, 0.78, 0.39, 0.195, and 0.098 μg/mL. The bacterial inocula were prepared by diluting (1:100) cultures grown in 10 mL Mueller–Hinton II broth for 6 h and the concentration adjusted to 0.5 McFarland turbidity standard. The antimicrobial susceptibilities were performed in 96-well microtiter plates (Becton and Dickinson). Plates were incubated at 37°C for 24 h and results were recorded as growth or no growth. Each concentration of the antibiotic was duplicated in the microtiter plate and MIC determinations were repeated with different inocula preparations.
Transferability of the tcrB gene
A conjugation assay to demonstrate transferability of tcrB gene from tcrB-positive to tcrB-negative isolates was performed by filter mating procedure (Tendolkar et al., 2006). The donor strains (14 tcrB-positive E. faecium and 1 E. faecalis) were resistant to tetracycline [tet(M) positive], erythromycin [erm(B) positive], and streptomycin (MIC ≥ 100 μg/mL) and susceptible to spectinomycin (mean MIC = 2.2 μg/mL). The E. faecium TX 5034 (Nannini et al., 2003) strain, resistant to spectinomycin (MIC > 100 μg/mL) and erythromycin [MIC ≥ 100 μg/mL; erm(B) positive] and susceptible to tetracycline [MIC = 0.78 μg/mL; negative for tet (M)], was used as the recipient for mating with tcrB-positive E. faecium isolates. The E. faecalis OG1SSp strain, resistant to streptomycin (MIC ≥ 100 μg/mL) and spectinomycin (MIC > 100 μg/mL) and susceptible to erythromycin [MIC = 0.48 μg/mL; negative for erm(B)] and tetracycline [MIC = 0.39 μg/mL; tet(M) negative], was used as the recipient for mating with tcrB-positive E. faecalis isolate. Transconjugants were selected on brain heart infusion (BHI) agar plates containing tetracycline (40 μg/mL) and spectinomcyin (500 μg/mL) for both E. faecium and E. faecalis isolates. Transconjugants were tested by PCR for tcrB, erm(B), and tet(M) genes. The MIC of copper for the two recipient strains and the 15 transconjugants were determined as described above. The transfer frequency for each isolate was calculated as the number of transconjugants per recipient CFU.
Pulsed-field gel electrophoresis
The pulsed-field gel electrophoresis (PFGE) analysis of tcrB-positive isolates was done as per Murray et al. (1990b) with minor modifications. Isolates were grown overnight in 5 mL of BHI broth at 37°C. The cells were pelleted by centrifugation and suspended in 0.85% NaCl. The plugs were prepared by mixing 200 μL of the cell suspension with 200 μL of 1.6% SeaKem gold agarose. The plugs were lysed by transferring them into a 10 mL lysis solution (6 mM Tris-HCl, pH 7.4, 100 mM ethylenediaminetetraacetic acid [EDTA], 1M NaCl, 0.5% sodium lauroyl sarcosine, 0.5% Brij, 0.2% deoxycholate, lysozyme [500 μg/mL], and RNaseA [20 μg/mL]) for 4 h at 37°C with gentle shaking. The plugs were then transferred to EDTA–sodium dodecyl sulfate–proteinase K (ESP) buffer (10 mM Tris-HCl, pH 7.4, 1 mM EDTA, 1% sodium dodecyl sulfate, and 50 μg/mL Proteinase K) and incubated overnight at 50°C with gentle shaking. The plugs were washed three times for 10 min each time with TE dilute buffer (10 mM Tris-HCl, pH 7.4, 0.1 mM EDTA) and stored at 4°C until used.
Restriction digestion of the plugs was performed with SmaI and the digested plugs were transferred onto a gel (1% SeaKem Gold Agarose with 0.5 × TBE) and placed in an electric field device, CHEF-DR II (BioRad®, Richmond, CA). The pulse time for block 1 was 3.5 s initial time and 25 s final time at 200 V for 12 h, and for block 2 was 1 s initial time and 5 s final time at 200 V for 8 h. Gel images were captured with a Gel Doc 2000 system (BioRad), and band patterns were analyzed and compared using BioNumerics software (Applied Maths, Austin, TX). We used the band-based Dice similarity coefficient and the unweighted pair group mathematical average algorithm method (UPGMA) for clustering with a position tolerance setting of 1.5% for optimization and position tolerance of 1.5% for band comparison. Isolates were grouped based on identical banding patterns (100% Dice similarity).
Statistical analysis
The prevalence of the tcrB gene in fecal enterococcal isolates was evaluated using a generalized mixed model (PROC GLIMMIX, SAS, Version 9.1.3). The statistical model included the fixed effect of dietary treatment and sampling week as a random effect. Because all isolates were resistant (MIC > 100 μg/mL) to erythromycin, oxytetracycline, and tetracycline, these data were not analyzed. The distribution of MIC values within copper or vancomycin and tcrB-negative or -positive isolates was evaluated for evidence of departure from normal distribution (PROC UNIVARIATE). Because the values were not normally distributed (p < 0.05), the values were transformed based on rank (PROC RANK), and analysis of variance (PROC GLIMMIX) was performed on the ranked values.
Nucleotide sequence accession number
The partial gene sequence of one tcrB-positive E. faecium (strain KSU-263) was deposited in the NCBI GenBank database (Accession number EU869871).
Results
Occurrence of the tcrB gene
A total of 323 enterococcal isolates were obtained from the feces of piglets collected at weekly intervals for 5 weeks (Table 2). All 323 enterococcal isolates were screened for the tcrB gene and 15 isolates (4.6%) were identified as positive (amplicon size was 663 bp) (Fig. 1). Sampling time (weekly) had no effect (p > 0.05) on the number of tcrB-positive enterococci isolated from the feces. Similarly, the number of tcrB-positive enterococci isolated was not different among the five dietary treatment groups (p > 0.05; Table 2). Seven tcrB-positive isolates (7/109; 6.4%) were from piglets fed an elevated supplemental level (125 ppm; copper or copper and zinc groups) and eight isolates (8/214; 3.7%) were from piglets fed a normal supplemental level (16.5 ppm) of copper in their diets (control, zinc, and antibiotic groups).
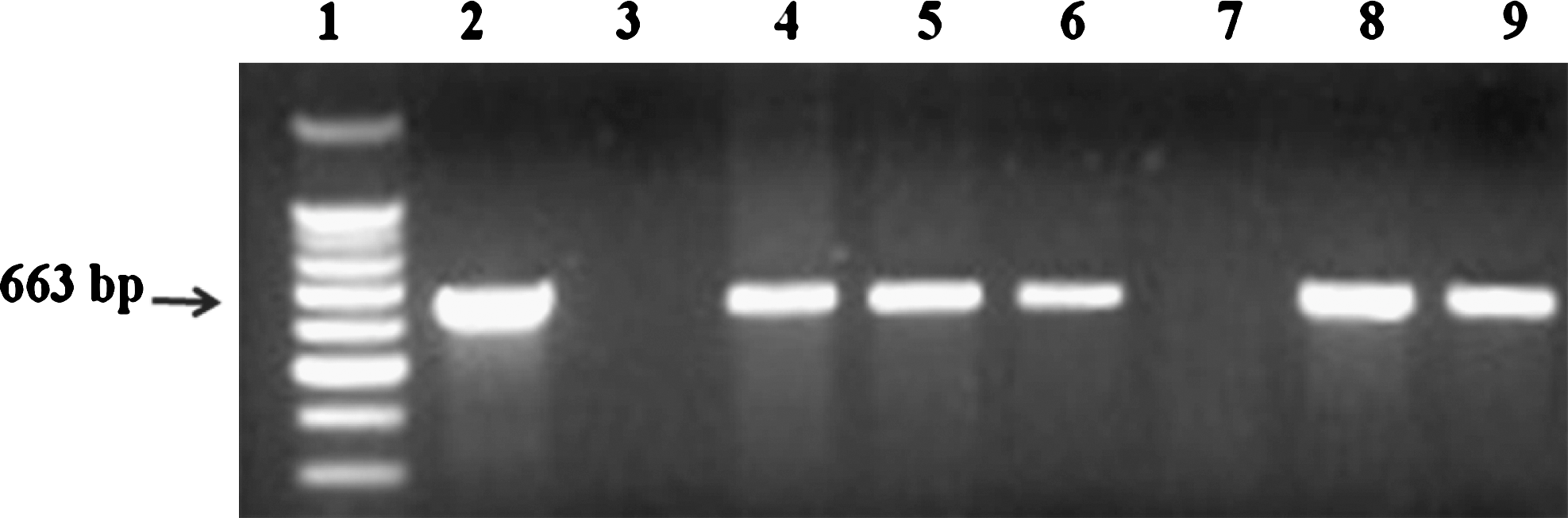
PCR amplification of the tcrB gene of enterococcal isolates from piglets fed diets supplemented with copper, zinc, copper and zinc, or neomycin and oxytetracycline. Lane 1: 100 bp molecular marker, Lane 2: Positive control (Strain from Denmark), Lane 3: Negative control, and Lanes 4–9: Isolates from this study.
Species identification
Based on API, the 15 tcrB-positive and the 15 randomly chosen tcrB-negative isolates were initially identified as E. faecium. The multiplex PCR revealed that one of the 15 tcrB-positive isolates, identified as E. faecium by API, was E. faecalis. The multiplex PCR confirmed the 15 tcrB-negative isolates as E. faecium. The sodA gene sequence analyses indicated that tcrB-positive or -negative E. faecium had 98%–99% sequence homology with the E. faecium and the tcrB-positive E. faecalis isolate had 99% homology with E. faecalis (data not shown).
Occurrence of erm(B), tet(M), vanA, and vanB
All 15 tcrB-positive and 15 tcrB-negative isolates were positive for the erm(B) and tet(M) genes and negative for the vanA and vanB genes.
MICs of copper
All tcrB-positive isolates obtained in our study (14 E. faecium and 1 E. faecalis) and the two Denmark strains of E. faecium grew on Mueller–Hinton agar containing copper at 16 or 20 mM. In contrast, the 15 tcrB-negative isolates were able to grow at 4 or 6 mM of copper concentration. The mean MIC of copper for tcrB-negative and tcrB-positive isolates were 6.1 (95% confidence interval = 2.21) and 21.1 mM (95% confidence interval = 4.68), respectively, and the difference was significant at p < 0.001. The two E. faecium strains, 7430162-6 and 7430275-4, from Denmark had a MIC of 23 and 24 mM, respectively.
MICs of antibiotics
All isolates (14 tcrB-positive E. faecium, 1 tcrB-positive E. faecalis, 2 tcrB-positive E. faecium from Denmark, and 15 tcrB-negative E. faecium) were resistant to tetracycline, chlortetracycline, erythromycin, and oxytetracycline with MIC greater than 100 μg/mL and susceptible to vancomycin. The MIC values for vancomycin were higher (p < 0.001) for tcrB-positive (0.39 μg/mL) than for tcrB-negative (0.098 μg/mL) isolates.
Transferability of the tcrB gene by conjugation
Fourteen tcrB-positive E. faecium isolates and 1 tcrB-positive E. faecalis were used to demonstrate transferability of the tcrB gene by conjugation. The 14 transconjugant E. faecium and 1 E. faecalis strains were positive for tcrB, erm(B), and tet(M) genes (Table 3). As expected, the transconjugants were resistant to tetracycline (MIC > 100 μg/mL) and erythromycin (MIC > 100 μg/mL) and were able to grow on BHI agar containing high copper concentrations (16 mM or higher). The mean MIC of copper for the 15 transconjugants was 17.6 mM. The mean transfer frequency for tcrB-positive E. faecium (14 isolates) was 1.01 × 10−5. The E. faecalis isolate had a transfer frequency of 1.16 × 10−5.
MIC, minimum inhibitory concentration.
PFGE of tcrB-positive enterococcal isolates
The clonal relationship among the 14 tcrB-positive E. faecium isolates was determined by PFGE typing. A total of six PFGE patterns (<95% Dice similarity) were observed among the 14 tcrB-positive E. faecium isolates (Fig. 2). Four of the five isolates from the zinc supplemented group had identical banding patterns (100% Dice similarity). Four of the five isolates from the copper- and zinc-supplemented group had different banding patterns (<95% dice similarity). The two Denmark isolates were clonally different and had distinctly different banding patterns compared to our tcrB-positive E. faecium isolates (Fig. 2).

Pulsed-field gel electrophoresis patterns of SmaI-digested genomic DNA of Enterococcus faecium isolates from piglets fed diets supplemented with none (control), copper, copper and zinc, or neomycin and oxytetracycline.
Discussion
Antibiotics are supplemented in swine diets, particularly of weanling and starter pigs, to reduce morbidity and mortality, and increase growth rate (Zhao et al., 2007). Similarly, copper, at elevated levels, is used as a dietary supplement in pigs for growth promotion (Cromwell et al., 1989; NRC, 1998). In piglets, the growth response to copper supplementation is in addition to the response provided by antibiotics (Edmonds et al., 1985). However, in European countries like Denmark, copper is included increasingly in swine diets at elevated levels, as a replacement to in-feed antibiotics, which have been banned for use as growth promotants (Hasman et al., 2006a). The mechanisms to explain the beneficial effects from copper supplementation are not fully known. It is suggested that copper exerts some systemic effect (Zhou et al., 1994) and more importantly beneficially alters gut microbial metabolism, which is because of the antimicrobial activity of copper (Højberg et al., 2005; Gould et al., 2009). The inhibition of potential pathogens, such as coliforms, and increased availability of nutrients and energy because of reduced microbial activity in the gut are contributing factors in growth promotion (Højberg et al., 2005).
In our study, only a small proportion of fecal enterococci (4.6%; 15/323), isolated from weaned piglets fed diets supplemented with a normal or elevated level of copper, contained the tcrB gene. Majority of the tcrB-positive isolates (14/15) were E. faecium and only one was E. faecalis. In Denmark, the tcrB was also detected in E. gallinarum, E. casseliflavus, and E. mundtii (Hasman et al., 2006b). The presence of the tcrB gene was associated with approximately fourfold increase (6 vs. 21 mM) in resistance to copper. Hasman and Aarestrup (2002) have reported a sevenfold increase (4 vs. 28 mM) in copper resistance in tcrB-positive E. faecium. For tcrB-negative isolates, we determined MIC of copper of only 15 E. faecium isolates that were randomly selected from 308 tcrB-negative enterococci. None of them had an MIC greater than 6 mM of copper. According to Hasman and Aarestrup (2002), all copper-resistant E. faecium strains isolated from feces harbored the tcrB gene and none of the copper susceptible isolates contained tcrB. Therefore, in enterococci, the presence of the tcrB gene is associated with resistance to copper.
In our study, the tcrB-positive enterococci were isolated from all five treatment groups, regardless of the copper level in the diet. Although we picked two esculin-positive colonies from each piglet sampled (three piglets per pen and six pens per treatment), we retained only one isolate after confirming the genus and species by esculin hydrolysis and the API and PCR analyses. This allowed us to screen a maximum of 18 isolates per sampling week per treatment group and the total isolates in each treatment group for the entire study ranged from 52 to 78. Interestingly, a higher proportion of enterococci positive for tcrB (7/109; 6.4%) were from groups supplemented with an elevated level (125 ppm) of copper (copper or copper and zinc group) compared to 3.7% (8/214) of isolates from groups fed a normal supplemental level (16.5 ppm) of copper in their diets (control, zinc, or antibiotic). Similarly, a higher proportion of enterococci positive for tcrB (10/121; 8.3%) were from groups supplemented with an elevated level (3000 ppm) of zinc (zinc or copper and zinc group) compared to 2.5% (5/202) of isolates from groups fed a normal supplemental level (165 ppm) of zinc in their diets (control, copper, or antibiotic). The influence of zinc supplementation on copper resistance is not fully known (Aarestrup and Hasman, 2004), although there is evidence that in Pseudomonas aeruginosa, copper exposure induces resistance to not only copper but also zinc (Caille et al., 2007). Of the seven tcrB-positive isolates in groups fed an elevated level of copper, six were E. faecium and they represented only four PFGE types based on SmaI restriction digestion. The clonal identities of the three isolates suggest possible pen-to-pen transmission. Of the eight tcrB-positive isolates from treatment groups fed a normal level of copper, only one isolate (from the antibiotic group) was clonally identical to an isolate (PFGE type 3) from the copper-supplemented group. The four tcrB-positive isolates from the zinc treatment group were from the same pen and had identical banding patterns (PFGE type 6), suggesting possible pig-to-pig transmission. Another explanation for the occurrence of the same clones may be the common source (same barn or litter mates) of piglets used in the study. Because of the limited number of isolates examined in the study, we could not ascertain whether an elevated copper level in the diet exerted pressure to select tcrB-positive enterococci. In testing different populations of E. faecium of animal or human origin in Denmark, Hasman and Aarestrup (2002) reported the highest level of copper resistance (76%) in isolates from pigs that had received 165 ppm of copper sulfate during the postweaning growth phase. In an in vivo animal challenge study (Hasman et al., 2006b), feeding 175 ppm of copper resulted in the selection of tcrB-mediated copper resistance in E. faecium compared to piglets fed low levels of copper (6 ppm).
An interesting aspect of copper resistance in enterococci is the genetic link of tcrB to macrolide and glycopeptide resistance (Hasman and Aarestrup, 2002). A 175-kb plasmid from a pig isolate of E. faecium that contained genes for vancomycin resistance (vanA) and macrolide resistance [erm(B)] along with the tcrB gene has been identified (Hasman and Aarestrup, 2002). In our study, all 15 tcrB-positive isolates contained the erm(B) gene and were phenotypically resistant to erythromycin (MIC = > 100 μg/mL). However, the tcrB-positive isolates did not contain either the vanA or vanB gene and were susceptible to vancomycin (MIC ≤ 0.7 μg/mL). Neither the presence of erm(B) nor the absence of van genes in enterococci is surprising. The erm(B) is the most common gene associated with enterococcal isolates resistant to macrolides (Jensen et al., 1999). Although erythromycin is not commonly used in swine, tylosin, another macrolide, is used in the feed to treat enteric bacterial infections and for growth promotion (Jackson et al., 2004b). We tested our isolates for both vanA and vanB. The vanB gene shares 76% of its amino acid sequence with vanA, and vanA is the most predominant resistant determinant in human enterococcal isolates in Europe, whereas vanB is most common in the United States (Cetinkaya et al., 2000). Occurrence of glycopeptide resistance in enterococci of pigs and chickens in Europe is was associated with the use of avoparcin, a glycopeptide, for growth promotion (Wegener et al., 1999; Aarestrup et al., 2001). However, the use of avoparcin in Europe was banned in 1997. Also, a genetic link between erm(B) and vanA has been documented in population of E. faecium isolates from pigs and chickens in Europe and Japan (Yoshimura et al., 1998; Aarestrup, 2000). The absence of glycopeptide resistance in enterococcal isolates of piglets in our study, which is in agreement with Poole et al. (2005), was possibly because avoparcin was never approved for use as a growth promotant in the United States (McDonald et al., 1997).
Enterococcal isolates in our study were also positive for tet(M) and phenotypically resistant to tetracyclines. There is evidence of co-selection of tet(M) with erm(B) in enterococci (Cauwerts et al., 2007). The tet(M) gene is most often carried by transposons of the Tn916-Tn1545 family, which are promiscuous conjugative transposons in both gram-negative and gram-positive bacteria (Chopra and Roberts, 2001). The carriage of tet(M) is associated with the erm(B) gene in Tn1545-like transposons (Clewell, 1995).
Antimicrobial-resistant enterococci are important opportunistic and nosocomial pathogens in humans (Moellering, 1992; Murray, 2000; Arias and Murray, 2008). Because of frequent occurrence of resistant enterococci in animals, the enterococci may serve as a reservoir of resistance genes capable of transferring to humans (Kojima et al., 2010). The medical importance of enterococci is related to the propensity of these organisms to participate in the horizontal transfer of the antibiotics and virulence genes. We were able to demonstrate the transferability of the tcrB gene from all strains of tcrB-positive enterococci isolated in our study. In case of E. faecalis, the recipient strain (OG1SSp) was negative for erm(B) and tet(M); therefore, we were able to demonstrate cotransfer of erm(B) and tet(M) with the tcrB gene. In case of E. faecium, we were able to demonstrate cotransfer of only tet(M) with the tcrB gene because the recipient strain was also positive for erm(B). Our results suggest that tcrB, erm(B), and tet(M) genes may be present on conjugative plasmids, but this remains to be determined.
An increased use of copper at elevated levels in swine feeds may exert selection pressure for copper resistance, mediated by the tcrB. Because of the genetic link between the tcrB gene and antibiotic resistance genes, copper supplementation may co-select for resistance to antibiotics, such as tetracyclines and macrolides. One of the reasons for persistence of macrolide and glycopeptides resistance in enterococci in the years following the ban of in-feed antibiotics in Europe may be the use of copper as a growth promotant (Boerlin et al., 2001; Hasman and Aarestrup, 2005).
Our study is the first report on the occurrence of the tcrB gene in enterococci of swine in the United States. The significance of this finding is the potential association between copper resistance and resistance to other antibiotics and the propensity of enterococci to transfer tcrB and antibiotic resistance genes to other strains within the same species and to other enterococcal species. Further studies are needed to determine whether supplementation with elevated levels of copper selects for copper resistance, and in turn co-selects for antibiotic resistance.
Footnotes
Acknowledgments
Contribution no. 10-051-J from the Kansas Agricultural Experiment Station. We thank Dr. Henrik Hasman, National Food Institute, Technical University of Denmark, for providing tcrB-positive strains, and Dr. Barbara E. Murray, Division of Infectious Diseases, University of Texas Medical School, for providing TX 5034 strain.
Disclosure statement
No competing financial interests exist.