
Abstract
Avian pathogenic Escherichia coli (APEC) is an important respiratory pathogen of poultry. A variety of virulence-associated genes and serogroups are associated with avian colibacillosis caused by APEC strains. One hundred forty-eight E. coli isolates recovered from diagnosed cases of avian colibacillosis from Guangdong province between 2005 and 2008 were serotyped, and characterized for virulence-associated genes, phylogenetic backgrounds, antibiotic susceptibility, and genetic relatedness. Associations between virulence-associated genes and antimicrobial resistance were further analyzed. Although 148 APEC isolates belonged to 21 different serogroups, 81% were of one of eight serogroups: O65 (27%), O78 (10%), O8 (9%), O120 (9%), O2 (7%), O92 (6%), O108 (5%), and O26 (5%). Polymerase chain reaction analysis showed that the most prevalent gene was traT (90%), followed by iroN (63%), fimH (58%), hlyF (55%), cvaC (54%), and sitA (51%). The APEC strains mainly belonged to groups A (73%) and D (14%). Multiple antimicrobial-resistant phenotypes (greater than or equal to three antimicrobials) were detected in all E. coli isolates, with the majority of isolates displaying resistance to tetracycline (97%), sulfamethoxazole (93%) and fluoroquinolones (87% for ciprofloxacin and 84% for enrofloxacin), chloramphenicol (74%), and florfenicol (66%). All E. coli isolates were further genetically characterized by pulsed-field gel electrophoresis. A total of 125 different pulsed-field gel electrophoresis profiles were obtained, implying that the multiresistant E. coli isolates carrying virulence-associated genes and belonging to multiple serogroups were not derived from a specific clone, but represented a wide variety of chromosomal backgrounds. Statistical analysis showed that several virulence-associated genes were significantly present in APEC isolates susceptibility to multiple antimicrobials. The findings demonstrate that a wide variety of serogroups and potential virulence genes, multiple-resistances, and the clear association of susceptibility and virulence genes have commonly emerged in APEC strains, and these also suggest that antimicrobials should be prudently used to reduce the emergence and spread of resistant strains carrying virulence-associated genes.
Introduction
E
Antimicrobials are commonly used for the treatment of avian colibacillosis caused by E. coli; however, increased resistance has been frequently reported in E. coli isolates from poultry in China (Yang et al., 2004; Zhao et al., 2005; Dai et al., 2008; Jiang et al., 2009). Resistant pathogens have emerged as a major concern for human and veterinary medicine. A study has reported the transmission of resistant E. coli clones from poultry to humans (Van den Bogaard et al., 2001). To tackle the development and spread of resistance and choose effective antimicrobial agents in various clinical settings, reliable data on the prevalence of resistance to specific antimicrobials of bacteria isolated from both humans and animals are required. Recently, a number of studies have reported that significantly reduced virulence potential was associated with resistance to certain antibiotics, such as fluoroquinolones, chloramphenicol, and tetracycline (Johnson et al., 2003; Horcajada et al., 2005; Moreno et al., 2006). However, up to now, the correlation between antimicrobials resistance and virulence-associated genes has not been studied in APEC strains isolated from south China.
The objective of this study was to determine the prevalence of serogroups, virulence-associated genes, phylogenetic groups, antimicrobial resistance, and genetic relatedness among APEC strains and to estimate the relationship of potential virulence genes with resistance to antimicrobial agents.
Materials and Methods
Bacterial strains
A large number of diseased or dead animals were submitted to the Veterinary Research Institute, Guangdong Academy of Agricultural Sciences, for diagnostic investigation. A total of 148 E. coli strains were isolated from broiler chickens found to have colibacillosis from November 2005 to March 2008. These diseased chickens were from 58 farms all over Guangdong Province, China. The farms typically housed ∼5000 animals. One to three flocks of chickens were sampled from each farm; each sample was from an individual animal. All samples were collected from a variety of tissues, including the air sac (2005, 5 isolates), trachea (2005, 15 isolates; 2007, 4 isolates; 2008, 11 isolates), liver (2005, 30 isolates; 2007, 14 isolates; 2008, 30 isolates), spleen (2007, 8 isolates), or heart blood samples (2005, 10 isolates; 2007, 18 isolates; 2008, 3 isolates) and inoculated onto MacConkey agar. The bacterial strains were identified by classical biochemical methods and confirmed by the API-20E system (bioMérieux). All strains were stored at −80°C in brain heart infusion broth with 15% glycerol until further use. For preparation of samples for testing, isolates were cultured on blood agar or trypticase-soy agar at 37°C over night.
Serotyping
The determination of O antisera was carried out by the method described by Orskov and Orskov (1990), by using antisera for all available somatic O types (O1-O171).
Virulence genotyping
All isolates were tested for 11 potential virulence genes by polymerase chain reaction (PCR) amplification. The primers used are listed in Table 1. Bacterial DNA was prepared by a rapid boiling method including a heating step at 100°C of a single colony in a volume of 100 μL of distilled water; after being cooled to room temperature, the suspension was centrifuged for 3 min at maximum speed to remove cell debris. Three multiplex PCR, used in the bulk of the genotyping studies, were as follows: (1) tsh, iss, and iucA; (2) hlyF and sitA; (3) ibeA, traT, cdtB, iroN, and cvaC. The fimH was detected in simple PCR assays. Amplification was done in a 25 μL volume reaction mixture containing 2 μL template DNA (except for 1 μL for fimH) and 50 pmol of each primer. All PCRs were done with the following conditions: 1 cycle of 5 min at 95°C; 30 cycles, each consisting of 30 sec at 95°C, 30 sec at annealing temperature, and 45 sec at 72°C; and 1 cycle of 5 min at 72°C. The PCR products were analyzed by electrophoresis and observed by ultraviolet transilluminator.
Phylogenetic analysis
Multiplex PCR-based method, as described by Clermont et al. (2000), was used for phylogenetic analysis. The primer pairs used were chuA, yjaA, and TSPE4.C2, which generate 279-, 211-, and 152-bp fragments, respectively (Table 1). The PCR was performed in a 25 μL volume consisting of 3 μL of template DNA and 50 pmol of each primer. The PCR steps were as follows: denaturation for 5 min at 94°C; 30 cycles of 30 sec at 94°C, 30 sec at 55°C, and 30 sec at 72°C, and a final extension step of 7 min at 72°C. The combinations of three genetic markers (chuA, yjaA, and TSPE4.C2) were used for determining the phylogenetic group of each strain (Clermont et al., 2000).
Antimicrobial susceptibility testing
All strains were determined for susceptibility to the following antimicrobial agents by agar dilution method: ceftriaxone, ceftiofur, amikacin, apramycin, streptomycin, gentamycin, chloramphenicol, florfenicol, tetracycline, doxycycline, sulfamethoxazole, ciprofloxacin, enrofloxacin, and levofloxacin. Antimicrobial susceptibility testing was conducted and the results were interpreted according to the guidelines of Clinical and Laboratory Standards Institute document M31-S1 (CLSI, 2004). As there are no CLSI breakpoints for doxycycline that are applicable to E. coli of animal origin, breakpoints of doxycycline (≥16 mg/L) were referred to CLSI document M100-S18 (CLSI, 2008) for isolates of human origin. E. coli ATCC 25922 was used as a quality control strain.
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) was used to analyze genomic relatedness among all APEC isolates. PFGE of chromosomal DNA digested with the restriction enzyme XbaI was carried out according to a standard protocol using a CHEF-MAPPER System (Bio-Rad Laboratories). Salmonella serovar Braenderup H9812 standards served as size markers. The gels were run at 6.0 V cm−1 with an angle of 120° at 14°C for 22 h. The results were interpreted according to the criteria described by Tenover et al. (1995).
Statistical analysis
Comparisons of the associations between resistance and potential virulence genes were carried out using Fisher's exact test (SAS, version 8.2; SAS Institute, Inc.). A p-value of <0.05 was considered significant; in such a case, odds ratio (OR) and their 95% confidence interval (CI) were calculated. For the purpose of this analysis, isolates with intermediate minimum inhibitory concentrations were considered susceptible.
Results
Serotyping
The majority of APEC strains (97%) were typeable with standard O antisera; for those that were typeable (146 isolates), a total of 21 different O serogroups were identified (Table 2). Among the isolates that could be typed, the most prevalent serogroups were O65 (27%), O78 (10%), O8 (9%), O120 (9%), O2 (7%), O92 (6%), O108 (5%), and O26 (5%). Other infrequent serogroups ranged from 1% to 3%, including O5, O53, O66, O79, O84, O88, O96, O115, O121, O123, O159, O160, and O163.
NT, nontypeable with standard O antisera.
Prevalence of virulence-associated genes
The prevalence of 11 virulence-associated genes was present in Table 2. The cdtB was not found in any of the E. coli isolates. None of the 11 virulence-associated genes was detected in two (1%) isolates. The traT was detected with the highest prevalence in all isolates. The iroN (63%), fimH (58%), hlyF (55%), cvaC (54%), and sitA (51%) genes were found in more than half of isolates. The distribution of virulence-associated genes related to serogroups showed that the traT gene was the most prevalent among typeable APEC isolates. The iroN and cvaC genes were detected in 79% of O8 strains. The genes for sitA and cvaC were found in 67% of O78- and O92-strains, respectively (Table 2). However, no significant association was found between 11 virulence-associated genes and O serogroups. With regard to seropathotypes, O78-sitA/iss/iroN/tsh/cvaC/traT (six isolates) was the most common, followed by O2-sitA/iss/iroN/hlyF/fimH/traT (four isolates) and O8-iss/iroN/hlyF/ibeA/cvaC/fimH/traT (four isolates) (data not shown).
Phylogenetic distribution of virulence-associated genes and serogroups
According to multi-PCR-based phylotyping, the majority of isolates belonged to phylogenetic group A (108 strains, 73%), followed by groups D (20 isolates, 14%) and B1 (18 isolates, 12%), whereas group B2 only accounted for 1% (2 isolates) of all isolates. Figure 1 displays the different profiles obtained by triplex PCR for the four phylogenetic groups. For all virulence-associated genes and serogroups considered, a very significant phylogenetic distribution emerged (Table 3). A significant association was observed between the presence of sitA and group D (p = 0.007, OR = 4.53; 95% CI, 1.44–14.30). A similar association was observed between the presence of tsh and D group (p < 0.001, OR = 5.32; 95% CI, 1.9–14.89). The differences in the distribution of O-serogroups with regard to phylogenetic groups were minimal, except for O92 (p = 0.019, OR = 6.15; 95% CI, 1.5–25.30) being significantly prevalent in group D than in other groups (Table 3).
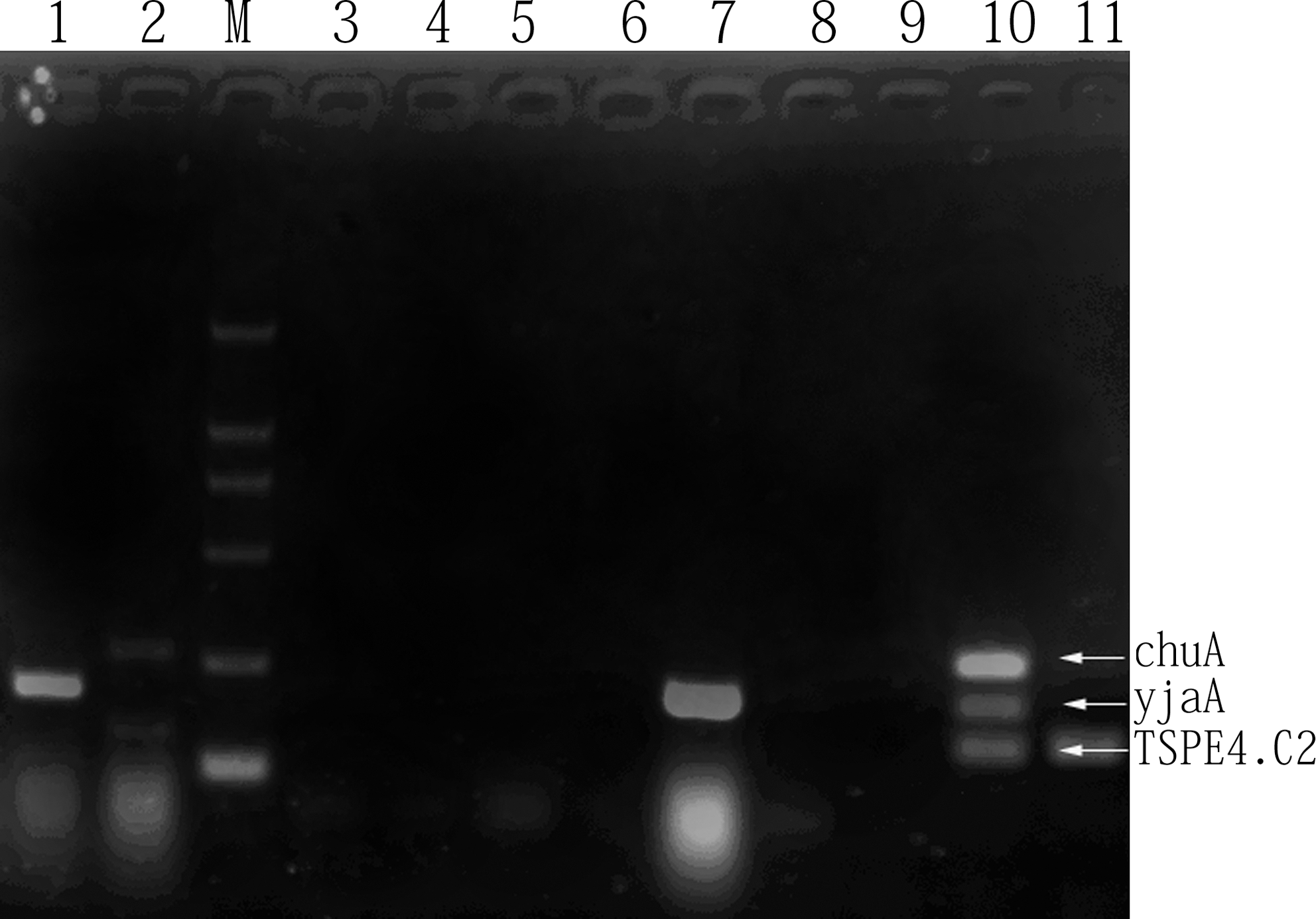
The polymerase chain reaction profiles specific for Escherichia coli phylogenetic groups. The combinations of chuA, yjaA, and TSPE4.C2 genes were used for determining the phylogenetic group of each strain. Lane 1, group A; lane 2, group D; lanes 3–9, group A; lane 10, group B2; lane 11, group B1; lane M, marker (100–2000 bp).
Only p-values <0.05 (by the Fisher exact test) are shown.
OR, odds ratio; CI, confidence interval.
Antimicrobial resistance phenotypes
The majority of the avian E. coli isolates were resistant to tetracycline (97%), sulfamethoxazole (93%), chloramphenicol (74%), and florfenicol (66%). Resistance to enrofloxacin and ciprofloxacin among APEC strains were quite high, which were found in 125 (84%) and 129 (87%) isolates, respectively, while to levofloxacin, in 51 (34%) isolates (Table 4). APEC isolates were less frequently resistant to apramycin (28%), ceftiofur (28%), ceftriaxone (19%), and amikacin (34%). With regard to multidrug resistance profiles, all isolates were resistant to >3 of the 14 antimicrobials tested, 40% were resistant to >10 antimicrobials, and 7% were resistant to >13 antimicrobials. Among multidrug resistance profiles, resistance to seven, eight, and nine antimicrobials were prevalent (17%, 18%, and 14%, respectively).
Association of antimicrobial resistance phenotype with virulence-associated genes
Of the 148 isolates, 143 were successfully typed by PFGE, and a total of 125 different PFGE profiles were obtained, suggesting that most of the isolates in the study were from epidemiologically unrelated E. coli clones. Possible statistical associations of antibiotic resistance/susceptibility phenotypes and virulence traits of epidemiologically unrelated isolates were further analyzed. However, the prevalence of resistance phenotypes according to phylogenetic groups was minimal (p > 0.05), except that enrofloxacin-susceptible isolates were significantly prevalent in phylogenetic group D (p = 0.02) (data not shown).
A more detailed analysis displayed associations of resistance/susceptibility phenotypes with potential virulence genes. First, none of virulence-associated genes were significantly associated with certain antimicrobial resistance/susceptibility phenotypes (including ceftriaxone, ceftiofur, gentamycin, tetracycline, ciprofloxacin, enrofloxacin, levofloxacin, florfenicol, and sulfamethoxazole); second, the presence of iss, cvaC, iroN/hlyF, tsh/cvaC, and iss/tsh was associated with susceptibility to amikacin, apramycin, streptomycin, chloramphenicol, and doxycycline, respectively (Table 5).
OR values above 1 represent positive associations and those below 1 represent negative associations.
Only the results for antimicrobial resistance that exhibited an association with virulence genes at a p < 0.05 are shown.
AMK, amikacin; APR, apramycin; STR, streptomycin; CHL, chloramphenicol; DOX, doxycycline; Vgs, virulence genes.
Discussion
Colibacillosis remains a major problem all over the world; however, it has not been established whether globally avian E. coli causing such infections share similar characteristics with regard to serogroups, virulence-associated genes, phylogenetic background, and antibiotic resistance. Therefore, the evaluation of these characteristics in avian E. coli from South China is very important.
Although a large number of serogroups have been described for E. coli strains, only a few of these, principally O1, O2, O78, O8, O35, and O36, have been commonly implicated in avian colibacillosis on a worldwide scale (Peighambari et al., 1995; Barnes et al., 2008). Of the eight predominant serogroups detected in this study, the prevalence of O2, O8, and O78 is in agreement with other studies (Blanco et al., 1998; Dho-Moulin and Fairbrother, 1999; La Ragione and Woodward, 2002; Ewers et al., 2004; Zhao et al., 2005), while the prevalence of O26, O65, O92, O108, and O120 is different from these reports. Unexpectedly, several studies found that other serogroups O4, O6, O8, O54, O61, O78, O88, and O93 were more prevalent in APEC isolates in the other geographic regions of China (Jin et al., 2008; Song et al., 2008). This discrepancy is interpreted by the fact that the distribution of serogroups can vary considerably from region to region. However, O35 and O36 serogroups reported by other people were not detected in any of the isolates. In addition to O2, O8, and O78 serogroups implicated in various human diseases, O26 detected in the study has been reported frequently associated with severe illness in humans (Brooks et al., 2005; Bielaszewska et al., 2007). Other infrequent serogroups, including O5, O53, O79, O88, O115, O123, O159, and O160 identified in this study, were also found in E. coli isolates from avian colibacillosis in other countries (Blanco et al., 1998; Monroy et al., 2005). However, several serogroups, including O26, O65, O84, O120, O121, O163, and O96, were not previously described in E. coli isolates from diseased poultry but in E. coli isolates from pigs (Gannon et al., 1988; Garabal et al., 1996; Leung et al., 2001; Fratamico et al., 2004). This supports a wide diversity of antigens present in APEC isolates, and may indicate that the emergence of the new serogroups associated with avian colibacillosis has not yet been reported.
The potential virulence genes, sitA, iss, tsh, iroN, hlyF, iucA, fimH, traT, and cvaC, were detected in all isolates in the study. These genes were also found in colibacillosis isolates from other countries (Janßen et al., 2001; Delicato et al., 2003; Rodriguez-Siek et al., 2005), which reveal that multiple potential virulence genes may participate in the pathogenesis of colibacillosis. However, the prevalence of these genes displayed relatively large differences in several studies. For instance, other studies showed that the tsh was detected in 19%, 49.7%, 84%, and 85.3% of the E. coli strains, respectively (Dozois et al., 2000; Janßen et al., 2001; de Brito et al., 2003; Zhao et al., 2005), whereas in 36% of the strains in this study and in 39.5% of the colibacillosis strains (Delicato et al., 2003). Similarly, the iss gene was detected in 83%, 86%, and 38.5% of the colibacillosis strains (de Brito et al., 2003; Delicato et al., 2003; Zhao et al., 2005), whereas it was found in 48% of the isolates in this study.
The fimH, cvaC, and traT genes were equally carried by E. coli isolates from cellulitis lesions and from feces (Peighambari et al., 1995; Ngeleka et al., 1996; de Brito et al., 2003); however, this could not be interpreted as a sign that these genes are unimportant in pathogenesis. For instance, fimH has been proven essential to human urinary tract infection (Mulvey et al., 1998), and F1 fimbriae are expressed by E. coli colonizing the trachea and the air sacs in vivo in chickens inoculated by the intratracheal route (Pourbakhsh et al., 1997), although other studies showed that fimH is not necessary for the colonization of trachea and air sacs (Marc et al., 1998; Arné et al., 2000). A study also suggests that these genes still might be required for infection (de Brito et al., 2003). It is plausible that different APEC strains can exploit several alternative paths to adhere, colonize, and invade their hosts.
Virulent extraintestinal pathogenic E. coli strains presumably belong mainly to group B2 and, to a lesser extent, to group D, whereas most commensal E. coli strains belong to group A (Clermont et al., 2000). However, phylogenetic analysis showed that APEC strains in this study mainly belonged to phylogenetic groups A and D. Several recent studies have reported similar results that APEC isolates frequently fall into A and D groups (Kariyawasam et al., 2007; Dissanayake et al., 2008; Ghanbarpour et al., 2010). This suggests that predictions about the virulence of APEC strains cannot be based only on chromosomal differences (Kariyawasam et al., 2007).
Antimicrobial therapy is one of the important measures for reducing significant economic losses to the poultry industry caused by colibacillosis. However, most of the APEC isolates from colibacillosis in this study showed high resistance to multiple antimicrobials such as tetracycline, sulfamethoxazole, and chloramphenicol, which also displayed multiple resistances to as many as five different antimicrobial classes. This observation is consistent with another Chinese study (Zhao et al., 2005), which is also similar to other reports (Sáenz et al., 2001; Gyles, 2008),
High resistance to doxycycline and florfenicol, two antibiotics now used legally and extensively in veterinary practice in China, was observed in this study; similar results were reported previously (Salehi and Bonab, 2006; Dai et al., 2008). The increase in resistance to doxycycline and florfenicol in this study is likely related to the increasing use of them on animal farms, or to the cross resistance as a result of the use of the same class such as tetracycline and chloramphenicol. The direct relationship between antimicrobial resistance and antimicrobial use was described by several studies (Moniri and Dastehgoli, 2007; Miranda et al., 2008; Varga et al., 2009).
Fluoroquinolones and cephalosporins are the most commonly used antimicrobials in both human and veterinary clinical medicine, but high resistance to fluoroquinolones has been reported: 90% of chicken isolates were resistant to ciprofloxacin (Sáenz et al., 2001) and more than half of all isolates were resistant to enrofloxacin and ciprofloxacin (Yang et al., 2004; Moniri and Dastehgoli, 2007). Likewise, in this study, most isolates also displayed high resistance to fluoroquinolones tested. Resistance to ceftiofur and ceftriaxone has increased over time, with the rates of 13% and 3% in 2000 (Zhao et al., 2005), 6.9% and 3.6% in 2003 (Liu et al., 2007), and 28% and 19% in the present study; thus, resistance to both antimicrobials should be carefully monitored in the future.
Physical linkages and statistical associations between resistance and virulence genes have been reported in swine pathogenic E. coli (Boerlin et al., 2005; Travis et al., 2006). However, in this study, we observed that some virulence-associated genes significantly present in APEC strains susceptible to amikacin, apramycin, streptomycin, chloramphenicol, and doxycycline, which are partly in agreement with previous reports that certain virulence-associated genes among human pathogenic E. coli isolates were significantly enriched in strains susceptible to certain antibiotics, such as chloramphenicol, tetracycline, and others (Johnson et al., 2003; Starčič Erjavec et al., 2007). This suggests that certain virulence genes may significantly exist in susceptible poultry as well as human strains. This indicates that the relationship of resistance with virulence-associated genes may vary according to particular host. The difference in such associations may be related to virulence-associated genes, geographical origin, and antimicrobial use of the strains under investigation.
Conclusions
These results reveal that a wide diversity of serogroups, multiple virulence-associated genes, and high resistance to multiple antimicrobial agents exist in avian E. coli isolates from colibacillosis. The analysis of genetic relatedness suggests that resistance has widely emerged among different APEC isolates. We believe that E. coli strains should be considered as important causative agents of development of avian disease, and antimicrobial resistance and virulence traits were as parts of an interconnected system. Virulence-associated genes significantly present in the susceptible APEC strains, suggesting that antimicrobial agents should be prudently used to control the emergence and increase of resistant isolates harboring virulence-associated genes.
Footnotes
Acknowledgments
We thank Professor Song Gao (College of Veterinary Medicine, Yangzhou University) for providing standard strains. This work was supported in part by the following grants: the National Natural Science Foundation of China (U0631006), Provincial Science Foundation of Guangdong (5200638), and the Program for Changjiang Scholars and Innovative Research Team in University (IRT0723).
Disclosure Statement
No competing financial interests exist.