
Abstract
Enterococcus faecium and Enterococcus faecalis belong to the gastrointestinal flora of humans and animals. Although normally regarded harmless commensals, enterococci may cause a range of different infections in humans, including urinary tract infections, sepsis, and endocarditis. The use of avoparcin, gentamicin, and virginiamycin for growth promotion and therapy in food animals has lead to the emergence of vancomycin- and gentamicin-resistant enterococci and quinupristin/dalfopristin-resistant E. faecium in animals and meat. This implies a potential risk for transfer of resistance genes or resistant bacteria from food animals to humans. The genes encoding resistance to vancomycin, gentamicin, and quinupristin/dalfopristin have been found in E. faecium of human and animal origin; meanwhile, certain clones of E. faecium are found more frequently in samples from human patients, while other clones predominate in certain animal species. This may suggest that antimicrobial-resistant E. faecium from animals could be regarded less hazardous to humans; however, due to their excellent ability to acquire and transfer resistance genes, E. faecium of animal origin may act as donors of antimicrobial resistance genes for other more virulent enterococci. For E. faecalis, the situation appears different, as similar clones of, for example, vancomycin- and gentamicin-resistant E. faecalis have been obtained from animals and from human patients. Continuous surveillance of antimicrobial resistance in enterococci from humans and animals is essential to follow trends and detect emerging resistance.
Introduction
Therapy of enterococcal infections may be difficult as enterococci are intrinsically resistant to a number of antimicrobial agents, including low-level resistance to beta-lactams, cephalosporins, clindamycin, and low-level resistance to aminoglycosides. Further, they can acquire resistance to other antimicrobial agents, including ciprofloxacin, erythromycin, tetracycline, linezolid, daptomycin, quinupristin/dalfopristin, and vancomycin (Murray, 1990; Arias and Murray, 2008). Invasive enterococcal infections are often treated with a combination of an aminoglycoside (e.g., gentamicin) and a cell-wall-active agent, such as penicillin (e.g., ampicillin) or a glycopeptide (vancomycin or teicoplanin), but due to emergence of resistant strains, new agents like linezolid, daptomycin, and quinupristin/dalfopristin are also used for therapy.
A nosocomial enterococcal infection can be regarded as a process in two stages. The first stage is an initial, usually asymptomatic colonization of the gastrointestinal tract by enterococcal strains. These strains can have different traits that may increase their survival in the patient, for example, antibiotic resistance, cytolytic toxin genes, or possible aggregation substances or other potential virulence genes. Subsequently, this population is expanded, often after the patient has been treated with antimicrobial agents eliminating a range of other competitive microorganisms in the intestine. In some patients, these intestinal enterococci may cause infections by tissue invasion either directly or indirectly (Mundy et al., 2000). In other cases, enterococcal infections are caused by nosocomial transmission of outbreak strains, which may not be related to the patient's own intestinal flora before hospitalization.
Enterococci can grow and survive in harsh environments and they have been isolated from soil, water, plants, most mammals, birds, and insects. E. faecium and E. faecalis are commensal bacteria in the intestine of humans and food animals, for example, poultry, cattle, and pigs. Contamination of meat with enterococci or other intestinal bacteria commonly occurs during slaughter of the animals. Large amounts of antimicrobial agents are used in animal production, and bacteria originating from food animals frequently carry resistance to a range of antimicrobial agents, including those regarded by the World Health Organization (WHO) as critically important for human therapy (WHO, 2007). Possible routes for transfer of either enterococci or resistance genes to hospitalized patients are shown in Figure 1.
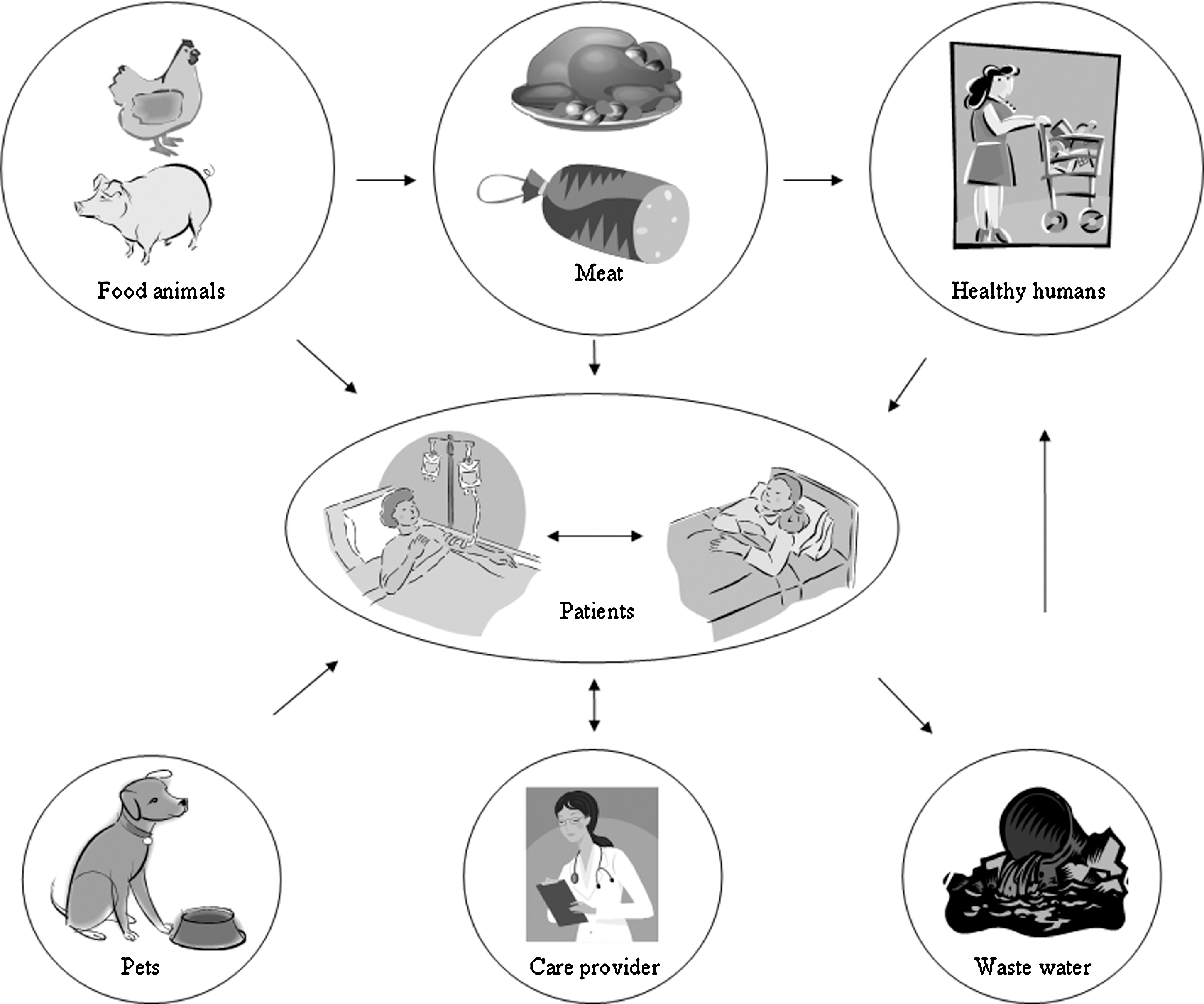
Possible routes for transfer of either enterococci or resistance genes to hospitalized patients.
This article describes the use of avoparcin, virginiamycin, and gentamicin in animals and the occurrence of antimicrobial resistance in enterococci in relation to this use. The finding of enterococci with similar resistance genes or similar genetic profiles obtained from animals, food, and humans are presented. Further, the article describes the transfer of resistance genes from enterococci of animal origin to enterococci of human origin.
Use of Antimicrobial Agents in Food Animals
In the 1940s and the early 1950s it was observed that the supplementation of different antimicrobial agents to feedstuff used for food animals improved the growth rate of animals. In the 1950s antimicrobial agents were added to commercial feed for poultry, cattle, and pigs in the United States (FDA, 2000), and this became a common practice in many parts of the world. Several antimicrobial agents used for growth promotion in animals are similar or identical to antimicrobial agents used in human therapy. This may constitute a human health hazard due to the risk of transfer of resistant bacteria or resistance genes from food animals to humans. In relation to treatment of enterococcal infections in humans, especially the use of the glycopeptides, avoparcin (analog to vancomycin) and the streptogramin, and virginiamycin (analog to quinupristin/dalfopristin) for growth promotion in food animals may imply a risk to human health (EC No 1831/2003). These antimicrobial agents are regarded by the WHO as critically important for human therapy (WHO, 2007). Another antimicrobial agent regarded as critically important for human therapy by WHO is gentamicin, which also is used in food animals.
Use of avoparcin in animals
The glycopeptide avoparcin was licensed in 1975 as an additive in feed used for food animals in many countries (although not in the United States and Canada). It has been used for broiler chickens, turkeys, pigs, beef and dairy cattle, calves, sheep, and goats. In 1993, vancomycin-resistant E. faecium were detected in farm animals in the United Kingdom (Bates et al., 1993). Studies performed in poultry flocks and pig herds fed with feedstuffs with or without avoparcin confirmed that avoparcin in the feed had a significant role in selecting for vancomycin-resistant E. faecium in the animals (Aarestrup, 1995; Bager et al., 1997). Since then, vanA, encoding vancomycin resistance, has been detected in E. faecium isolates from pets and farm animals, environmental samples, sewage, meat, stool samples from farmers, and nonhospitalized humans in the community from many parts of the world (Bates et al., 1994; Torres et al., 1994; Klare et al., 1995; Devriese et al., 1996; van Belkum et al., 1996; Werner et al., 1997; Wegener et al., 1997; Simonsen et al., 1998; Descheemaeker et al., 1999; Robredo et al., 2000; Guardabassi et al., 2002; Hammerum et al., 2004; Novais et al., 2005; Messi et al., 2006; Jung et al., 2007; Arriola et al., 2008). Subsequently, the use of avoparcin was banned in Denmark and Norway in 1995, Germany in 1996, and in the rest of the European Union (EU) in 1997, based on the precautionary principle due to the concern for human health. Avoparcin was banned in Korea in 1997 and in Taiwan and New Zealand in 2000 (Manson et al., 2003; Lim et al., 2006a; Lauderdale et al., 2007). In Australia, the use of avoparcin ceased in June 2000 (J. Suter, pers. comm.). To our best knowledge, the use of avoparcin is by now also discontinued in many Latin American countries (A. Vieira, pers. comm.).
Recently, vancomycin-resistant E. faecium were detected in feces from pigs in the United States. The origin of these vancomycin-resistant E. faecium isolates is notable since avoparcin was never used for pigs or other animals in the United States (Donabedian et al., 2009).
Since 2000, vancomycin-resistant E. faecium have been detected in poultry in Sweden, even though avoparcin has not been used since the early 1980s (Nilsson et al., 2009). Several studies have shown that vancomycin-resistant E. faecium persisted in animals and humans for an extended time period after the ban of avoparcin and persistence in the animal environment was demonstrated (Borgen et al., 2000; Heuer et al., 2002a, 2002b; Hammerum et al., 2004; Novais et al., 2005; Lim et al., 2006a; Sørum et al., 2006; Garcia-Migura et al., 2007; Ghidan et al., 2008).
The persistence of vancomycin resistance in E. faecium may have several explanations. In Denmark, vancomycin-resistant E. faecium persisted in pigs at a frequency around 20% for some time after the ban of avoparcin in 1995 (Fig. 2). It was not until the cessation of use of another antimicrobial agent (tylosin—a macrolide also used for growth-promotion) that the occurrence of vancomycin-resistant E. faecium decreased to only a few percent (Aarestrup et al., 2000). This was explained by coselection since the erm(B) gene encoding tylosin resistance and the vanA gene were located close to each other on the same plasmid (Aarestrup, 2000). Another possible explanation for the persistence of vancomycin-resistant E. faecium isolates is a link between Tn1546 and the copper resistance gene tcrB. Copper sulfate is used as a growth-promoting feed supplement for food animals. Hasman et al. showed a correlation between copper resistance and resistance to macrolides and glycopeptides as the tcrB gene was found on a plasmid containing both vanA and erm(B). However, the continued use of copper sulfate after the ban of avoparcin has not caused continued high levels of glycopeptide resistance, indicating that coselection might occur but not at a high level (Hasman and Aarestrup, 2005a; Hasman et al., 2005b). A third explanation is that vanA is linked with postsegregational killing systems on mobile genetic elements. These systems encode an unstable antitoxin that neutralizes a stable toxin encoded by the same plasmid. Antitoxin must be produced continuously to prevent the longer lived toxin from killing the cell. If vancomycin resistance is linked to such a plasmid, this mechanism creates a replication advantage for plasmid containing bacteria. Different classes of postsegregational killing systems linked to vanA have been described: mazEF, axe-txe, and ω-ɛ-ζ (Grady and Hayes, 2003; Moritz and Hergenrother, 2007; Sletvold et al., 2007). E. faecium isolates with a plasmid harboring the vanA transposon Tn1546 linked to the postsegregational killing system ω-ɛ-ζ has been shown to be stable in the intestine of mice for >20 days in the absence of vancomycin (Dahl et al., 2007).
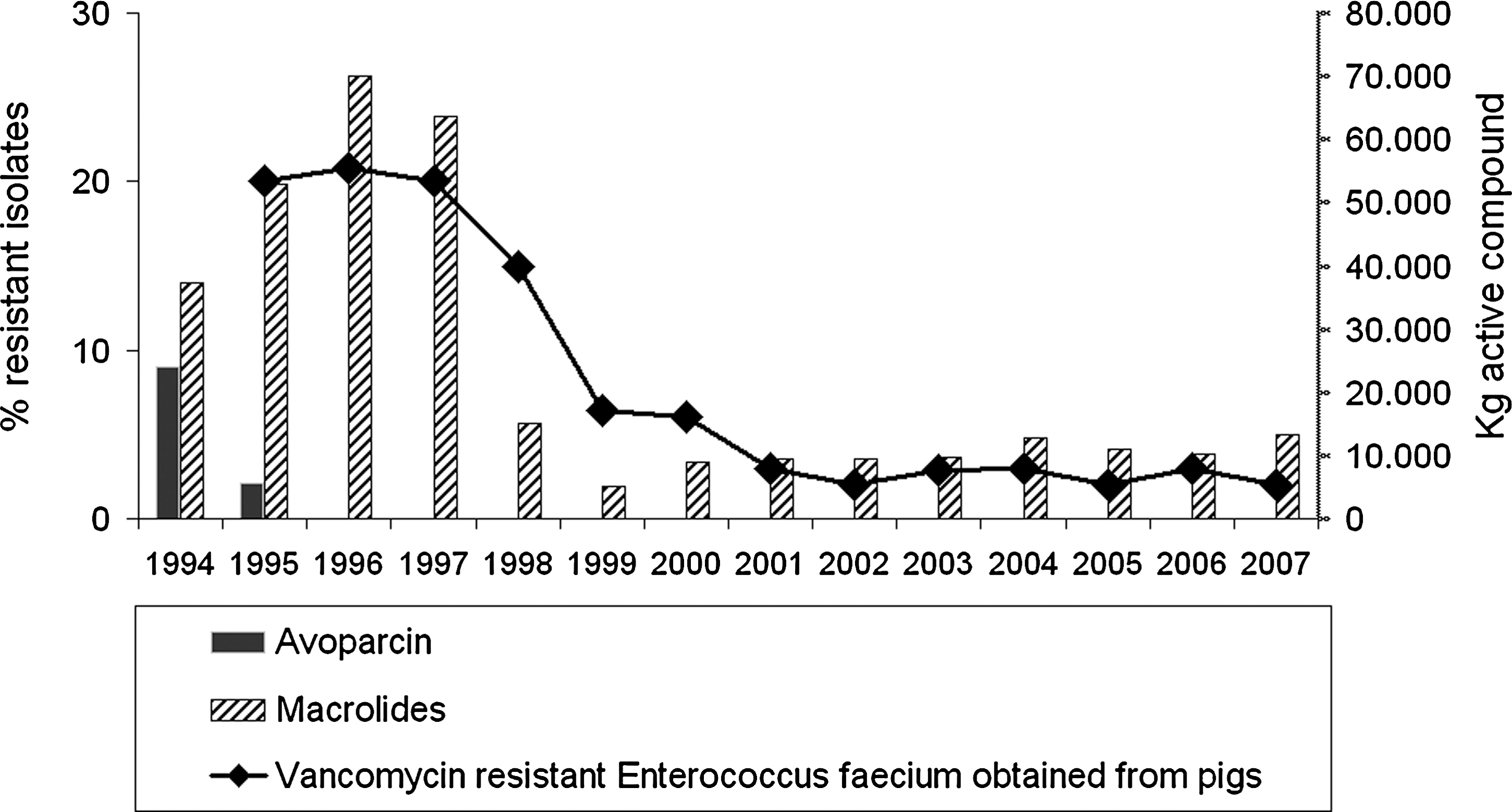
Occurrence of vancomycin resistance among Enterococcus faecium isolates from fecal samples from pigs (1995–2007) and consumption of avoparcin and macrolides (tylosin). (Data obtained from DANMAP 2007, 2008.)
Use of virginiamycin in animals
The streptogramin complex, virginiamycin, has been widely used as a feed additive in several countries, including the United States, where avoparcin was never used. In the EU, virginiamycin was used as a growth promoter in turkeys, chickens, laying hens, cattle, piglets, and pigs. Virginiamycin has also been used to prevent necrotic enteritis and coccidiosis in chickens and to control swine dysentery.
In 1997, quinupristin/dalfopristin-resistant E. faecium were isolated from nonhospitalized humans in the United Kingdom (Woodford et al., 1997). This finding was rather surprising since the new streptogramin, quinupristin/dalfopristin, used for human therapy, was not licensed in the United Kingdom at that time for use in humans. In studies from the United States and Europe, findings of quinupristin/dalfopristin-resistant E. faecium isolates outside the hospitals were assumed to be associated with use of virginiamycin for animals (Welton et al., 1998; Aarestrup, 2000).
In 1997, only one gene, satA [renamed vat(D)], was known to encode streptogramin A resistance in E. faecium. In 1998, satA was detected in E. faecium from poultry, pigs, pork, sewage, and animal manure (Hammerum et al., 1998; Jensen et al., 1998b; Werner et al., 1998). In 1999, a new gene, satG [renamed vat(E)], encoding streptogramin A resistance in E. faecium was described (Werner and Witte, 1999). vat(E) was detected in isolates from sewage, broiler chickens, and patients in different European countries and the United States (Werner and Witte, 1999; Soltani et al., 2001; Donabedian et al., 2006). Virginiamycin was banned in Denmark in 1998 and in all of EU in 1999. In Australia, the use was restricted to therapeutic purposes in 2008 (J. Suter, pers. comm.). Streptogramin resistance in isolates from animals persisted several years after the ban of virginiamycin (de Jong et al., 2009; Hammerum et al., 2009). However, data on the occurrence of streptogtamin resistance should be interpreted with care as different breakpoints for quinupristin/dalfopristin have been used. This may have resulted in reporting of a more frequent occurrence of streptogramin resistance among food animals, when using the Clinical and Laboratory Standards Institute breakpoint (≥4 μg/mL) compared with the use of the European Committee on Antimicrobial Susceptibility Testing breakpoint (>4 μg/mL) (Hammerum et al., 2009).
Use of gentamicin in animals
Gentamicin is commonly used in swine and widely used in chickens and turkeys in the United States (Donabedian et al., 2003). In the EU, gentamicin has not been used as a growth promoter, but it is used for therapy in food animals and in companion animals. In 1998, high-level gentamicin-resistant E. faecalis was reported in fecal samples from pigs in Denmark (DANMAP 97, 1998). In 2001, Donabedian et al. (2001) reported frequent isolation of high-level gentamicin resistance enterococci in food animals from the United States. Later high-level gentamicin resistance in enterococci obtained from poultry, pigs, dogs, and healthy humans was reported in several studies from other countries, (Lim et al., 2006b; Fracalanzza et al., 2007; Chan et al., 2008; Abbott et al., 2009; Jackson et al., 2009).
High-level gentamicin resistance in enterococci from infections in human is most often encoded by aac(6′)-Ie-aph (2″)-Ia, which encodes the bifunctional enzyme AAC(6′)-APH(2″). In 2003, aac(6′)-Ie-aph (2″)-Ia was also detected in E. faecium and E. faecalis isolates obtained from meat and food animals in the United States (Donabedian et al., 2003). Later, aac(6′)-Ie-aph (2″)-Ia was found in enterococci obtained from poultry in Malaysia and in E. faecalis from Danish pork, healthy pigs, nonhospitalized humans, and from patients with endocarditis (Chan et al., 2008; Larsen et al., unpublished data).
In conclusion, vancomycin-resistant enterococci, quinupristin/dalfopristin-resistant E. faecium, and gentamicin-resistant enterococci have been isolated from food of animal origin and from fecal samples from food animals. The findings of resistance to vancomycin and quinupristin/dalfopristin in enterococci of animal origin have been ascribed to the use of these antimicrobial agents as growth promoters in animal feed. The use of avoparcin is currently discontinued or reduced in many parts of the world. The use of virginiamycin was discontinued in Europe in 1999, but still in use for growth promotion and therapy in other parts of the world. Data on the use of gentamicin for food animals is limited; however, gentamicin-resistant enterococci are reported in several food animal species.
In many countries, interventions have led to significant reductions of nontherapeutic use of vancomycin and virginiamycin in food animals, and subsequent reductions in the occurrence of resistance to these antimicrobial agents in isolates from food animals. However, at the same time, enterococci resistant to these antimicrobial agents seem to persist in animals and in the environment several years after discontinuation of use.
Transfer of Enterococci Between Animals and Humans
Several studies have addressed the possible exchange of antimicrobial-resistant enterococci between animals and humans. Some of theses studies are presented below.
Comparison of E. faecium isolates obtained from animals and humans
Molecular typing of E. faecium isolates obtained from different sources has been used to investigate potential associations between the use of antimicrobial growth promoters and the risks to human health. During the last decade of the 20th century, pulsed-field gel electrophoresis (PFGE) was the golden standard for investigation of enterococcal outbreaks in hospitals, and this method was also applied for comparison of E. faecium isolates of animal origin with E. faecium isolates of human origin. Similar and highly similar PFGE profiles were found for vancomycin-resistant animal isolates and human stool isolates (van den Bogaard et al., 1997; Jensen et al., 1999; Hammerum et al., 2004).
In 2000, amplified-fragment length polymorphism analysis was used to investigate the genetic relationships among 255 vancomycin-resistant E. faecium strains isolated from hospitalized patients, nonhospitalized humans, and various animal sources. Four major amplified-fragment length polymorphism genogroups (A–D) were discriminated. Genogroup A consisted of isolates from pigs and nonhospitalized humans. Almost all isolates from poultry belonged to genogroup B, whereas most of the hospitalized patients belonged to genogroup C. Genogroup D consisted of isolates from veal calves and veal calf farmers. This study lead to the conclusion that vancomycin-resistant E. faecium strains were predominantly host specific, and strains isolated from hospitalized patients were genetically different from the prevailing vancomycin-resistant E. faecium strains present in the fecal flora of nonhospitalized humans (Willems et al., 2000).
In 2002, multilocus sequence typing (MLST) was developed for E. faecium. In the first study, internal fragments from 7 housekeeping genes of 123 epidemiologically unrelated isolates from humans and livestock in The Netherlands and 16 human-derived isolates from several outbreaks in the United States, the United Kingdom, Australia, and The Netherlands were analyzed. Outbreak isolates from hospitalized humans clustered in a subgroup, named clonal complex 17 (CC17) (Homan et al., 2002). Other studies showed that CC17 was associated with hospital-related isolates and was not found among E. faecium isolates of animal origin (Top et al., 2004; Willems et al., 2005). E. faecium isolates belonging to CC17 are ampicillin resistant and high-level ciprofloxacin resistant, and many are vancomycin resistant and carry specific virulence genes (Leavis et al., 2006a, 2006b). Human outbreak E. faecium isolates belonging to CC17 has been found in at least five continents, and molecular evidence of host specificity was brought forward (Willems et al., 2000; Leavis et al., 2006a). Galloway-Pena et al. (2009) looked at 53 hospital-outbreak E. faecium isolates from the United States from 1970 to 1991. The earliest CC17 isolate in there study was from 1982. This could indicate that CC17 isolates have been hospital-adapted during the last decades. In the same study, strains from some of the outbreaks from the 1970s and 1980s belonged to CC9, which has been associated with poultry.
Later studies have questioned the host specificity of CC17. Studies have shown that dogs can be a reservoir of E. faecium isolates belonging to sequence type 78, which is part of CC17 E. faecium complex (Damborg et al., 2008, 2009). In another study vancomycin-resistant E. faecium isolates with sequence type 132 (ST132 is part of CC17) were obtained from a pig and from a human urinary tract infection from Portugal (Freitas et al., 2009b). Further, the vanB2 gene has been detected in an E. faecium isolate from chicken meat and an E. faecium isolate from veal in a Spanish study. Both of these isolates were ST17, which is the most common sequence type in CC17 (Lopez et al., 2009). In the same study, a vanA-positive E. faecium isolate was obtained from rabbit meat; this isolate was ST78, which also is a part of CC17 (Lopez et al., 2009).
Most often, E. faecium isolates from pigs belong to one cluster (CC5) by MLST. E. faecium belonging to CC5 (ST5, ST6, and ST185) have been found in pigs from five European countries (Homan et al., 2002; Freitas et al., 2009b; A.M. Hammerum, unpublished data). Isolates belonging to CC5 have also been found in patients with urinary tract infections and in fecal samples from nonhospitalized humans (Freitas et al., 2009b; A.M. Hammerum, unpublished data).
Comparison of E. faecalis isolates obtained from animals and humans
MLST of E. faecalis have shown several clonal complexes (CC2, CC9, and CC87) related to outbreaks in hospitals (Ruiz-Garbajosa et al., 2006; Kawalec et al., 2007; Freitas et al., 2009a). In a recent study an E. faecalis isolate with ST6 (CC2) was obtained from a pig sample from Portugal (Freitas et al., 2009a). E. faecalis isolates with ST6 have previously been found in patients from hospitals in Poland, Portugal, Spain, and Italy and from healthy Norwegian infants (Kawalec et al., 2007; Freitas et al., 2009a, 2009b; Solheim et al., 2009).
E. faecalis isolates classified as ST16 have been obtained from pigs, healthy humans, and poultry in Portugal and Spain; from a healthy Norwegian infant; and from patients in Poland (Ruiz-Garbajosa et al., 2006; Kawalec et al., 2007; Freitas et al., 2009a; Solheim et al., 2009).
Recently, high-level gentamicin-resistant E. faecalis belonging to ST16, and with similar PFGE types, have been obtained from pigs, pork, nonhospitalized humans, and patients with endocarditis (Larsen et al., 2010). The pig isolates in the study also contained part of an enterococcal pathogenicity island (Shankar et al., 2006). In another study, the same sequence type, ST116, was found in vanA-positive E. faecalis isolates from turkey meat, nonhospitalized humans, and a patient in Denmark (Agersø et al., 2008). Another study has shown that sensitive E. faecalis isolates obtained from patients with endocarditis had the same sequence types (ST19, ST21, and ST72) as E. faecalis isolates obtained from nonhospitalized humans (J. Larsen, pers. comm.). Larsen et al. also found E. faecalis of ST40 and ST97 from pigs and from endocarditis patients (J. Larsen, pers. comm.).
In conclusion, E. faecium isolates obtained from hospital outbreaks are associated with CC17. E. faecium isolates of animal origin most often have other sequences types, even though isolates belonging to CC17 have been found among strains of animal origin. Host specificity is less evident among E. faecalis, and the same sequence types have been detected in E. faecalis of human and animal origin. However, some E. faecalis clonal complexes are considered high-risk clonal complexes (CC2, CC9, CC87) as they mainly consist of isolates causing infections in humans.
Gene Transfer Between Enterococci of Different Origin
In several studies, investigators have looked for a link between genetic elements in enterococci of human and animal origin. Two studies have found similar plasmids obtained from animals and humans, indicating gene transfer between different hosts (Lim et al., 2006b; Sletvold et al., 2007). Most other studies have characterized Tn1546, the transposon on which vanA is located. Other studies have looked at transfer of resistance genes from enterococci of animal origin to enterococci of human origin in vitro and in vivo. Several of the studies are described below.
Characterization of Tn1546
The vanA resistance gene is located on the transposon Tn1546 and was first described in an E. faecium clinical isolate by Arthur et al. (1993). Tn1546 consists of 10,851 base pairs and encodes 9 polypeptides that can be assigned to various functional groups: transposition (ORF1 and ORF2), regulation of resistance gene expression (VanR and VanS), synthesis of the D-Ala-D-Lac depsipeptide (VanH and VanA), and hydrolysis of peptidoglycan precursors (VanX and VanY); the function of VanZ remains unknown.
Jensen found a point mutation in the vanX gene (G to T) at position 8234 in Tn1546. The mutation to T has been associated with a pig origin of the vancomycin E. faecium isolates, whereas a G in position 8234 is found in vancomycin-resistant E. faecium isolates of poultry origin. Both types can be found among E. faecium isolates of human origin (Jensen, 1998; Novais et al., 2008). Woodford et al. (1998) studied the diversity of vanA elements in 64 human enterococci from patients at 15 hospitals in the United Kingdom and compared these with 42 enterococci of nonhuman origin from the United Kingdom. The isolates were divided into 24 groups by polymerase chain reaction analysis of vanA elements. Even though the vanA elements in the studied enterococci were heterogeneous, six groups were shared by enterococci of human and nonhuman origin.
Novais et al. (2008) studied the diversity of Tn1546 in 176 Portuguese enterococci of human and nonhuman origin from 1996 to 2004. As for the United Kingdom isolates, high diversity has been observed among the Portuguese isolates; 24 Tn1546 variants were detected (all types were plasmid borne). Three types were only associated with farm animals, 11 types with hospital patients, and 1 type with healthy human volunteers, whereas 8 types were recovered from different sources. Molecular diversity and evolutionary relationship of Tn1546 were also studied by Willems et al. (1999). The studied collection consisted of isolates of both human and animal origin and was collected from The Netherlands, the United States, and the United Kingdom. As in the other studies, Willems et al. found a great diversity among the isolates, but they found identical Tn1546 types in isolates from humans and farm animals in The Netherlands. Similar findings were observed in a study of Danish vancomycin-resistant E. faecium isolates. Here, Jensen et al. (1998a) also found E. faecium isolates with similar Tn1546 elements from humans and animals, even though isolates with several different Tn1546 types were investigated. Tn1546 from animal, food, and human E. faecium isolates were compared by Biavasco et al. (2007) to study possible reservoirs and transmission routes. They found that Tn1546 was plasmid borne in most (95%) of the studied isolates and most of the isolates had the Tn1546 prototype. In this study, no significant associations between Tn1546 type and the source of the vancomycin-resistant enterococcal isolate were found (Biavasco et al., 2007).
In conclusion, several studies have shown many different variants of Tn1546 among vancomycin-resistant enterococci. However, the same types have been found in enterococci of human and animal origin. Further, Tn1546 are often plasmid borne. Taken together, this indicates horizontal gene transfer of vanA between enterococci of animal and human origin.
Animal models for gene transfer studies between enterococci
Enterococci can harbor different transferable genetic elements such as conjugative transposons and different types of plasmids with resistance genes. An ideal place for gene transfer could be the gut of different animal species, including humans. Different animal models have been used to test this hypothesis.
Antibiotic administration is known to increase the susceptibility of the host to colonization with antibiotic-resistant bacteria. A model was developed by Hentges et al. (1985); they administered five different antimicrobial agents (ampicillin, clindamycin, kanamycin, metronidazole, and streptomycin) in the drinking water to mice and found that streptomycin was the most effective antimicrobial agent to decrease the colonization resistance. Streptomycin is an aminoglycoside that in vitro has no effect on anaerobic bacteria. However, administration of 5 g/L streptomycin in the drinking water of mice disrupts both aerobic and anaerobic components of the intestine (Hentges, 1984).
The model has been used to study colonization of E. faecium strains and transfer of resistance genes between E. faecium strains (Whitman et al., 1996; Lester et al., 2004).
Conjugal transfer of resistance genes between enterococci in gnotobiotic animals has previously been described (Jacobsen et al., 1999; Moubareck et al., 2003; Dahl et al., 2007). In these studies the level of recipients, donors, and transconjugants was much higher than that obtained in streptomycin-treated mice. An explanation for the observed difference may be explained by the use of gnotobiotic animals. Gnotobiotic animals have no colonization resistance, whereas streptomycin-treated mice still have some colonization resistance, even though it is decreased compared with untreated animals.
Several other in vivo gene transfer models for enterococci have been described. Transfer of a tetracycline resistance gene has been shown in streptomycin-treated mini-pigs (Licht et al., 2002,) and transfer of the vanA gene cluster has been shown in mice harboring a human microbiota (Mater et al., 2005; Bourgeois-Nicolaos et al., 2006). In vivo transfer rates in the intestine may be higher than assessed in vitro in the laboratory (Dahl et al., 2007).
The different models have shown that the intestine is a hot spot for transfer of resistance genes between enterococci. Gnotobiotic animal models are far from the natural situation happening in humans. The best animal model imitating the situation in human patients would be one testing gene transfer in animals treated with an antimicrobial agent often used for treatment of human clinical infection (e.g., cephalosporins and flouroquinolones).
Colonization and gene transfer studies with enterococci in the human intestine
Transient colonization of enterococci of animal origin has been shown in the intestine of humans. A study by Berchieri showed colonization of two vancomycin-resistant E. faecium isolates: one isolated from a chicken and the other isolated from a pig. The chicken isolate colonized the intestine for 20 days, whereas the pig isolates colonized the intestine for only 4 days (Berchieri, 1999). However, this experiment was preformed on only one subject and it was the same subject who ingested first the isolate of chicken origin and then 25 days after the isolate of pig origin. Sørensen et al. (2001) showed transient colonization in 12 subjects of vancomycin- or streptogramin-resistant E. faecium isolates of animal origin. The subjects were colonized for approximately 1 week. However, none of these studies were designed to investigate the possible transfer of resistance genes.
Previous studies have also shown transient colonization of the intestine after ingestion of enterococci-contaminated food (Sørensen et al., 2001; Lund et al., 2002; Gelsomino et al., 2003).
Transfer of resistance genes in the human intestine was tested in a study by Lester et al. (2006). This study demonstrated transfer of the vanA gene cluster between an E. faecium isolate of animal origin to an E. faecium isolate of human origin in the intestine of humans. In vivo conjugation occurred in three out of six volunteers. These three volunteers were all heavily colonized with both the donor and the recipient strain. Conversely, the other three volunteers showed lower maximum concentrations or a delayed increase in concentration of the strains. This indicates that for transfer to occur, high numbers of both the donor and the recipient strains must be present at the same time. In one volunteer cotransfer of several resistance genes occurred. The vanA gene was transferred together with the vat(E) gene and the erm(B) gene (encoding streptogramin and macrolide resistance).
The recipient strain in this study did not belong to CC17, but a recent study has shown that vanA of animal origin can be transferred to a CC17 recipient obtained from a patient with sepsis. This study was performed in the intestine of cephalosporin-treated mice (C.H. Lester, unpublished data).
In conclusion, the different studies of transposons, plasmids, and of gene transfer indicate that even though there may be some degree of host specificity among enterococci, resistance genes can be exchanged between enterococci from different reservoirs, and thereby antimicrobial-resistant enterococci of animal origin may constitute a hazard to human health.
Conclusions
Several studies have shown that genes encoding vancomycin, quinupristin/dalfopristin, or gentamicin resistance are similar in E. faecium isolates obtained from human patients and animals/meat. Most E. faecium causing outbreaks belong to CC17, whereas E. faecium isolates from animals and humans in the community most often belong to other clonal complexes. The findings of the same genes in different host indicate gene transfer from E. faecium of animal origin to E. faecium isolates of human origin. This gene transfer may happen in the intestine of human patients, from an E. faecium strain of animal origin (obtained before the hospital stay or from eating meat containing resistant E. faecium strains during the stay) to a hospital CC17 E. faecium strain obtained during the hospital stay.
E. faecium from production animals or meat are therefore most often not a human hazard in themselves, but E. faecium of animal origin can act as a donor of antimicrobial resistance genes for other pathogenic enterococci. Further, new studies have shown that E. faecium from dogs can belong to CC17, indicating that dogs can be a reservoir of pathogenic E. faecium.
The situation is different for E. faecalis. The same clonal complexes of vancomycin-resistant, gentamicin-resistant, and sensitive E. faecalis isolates have been obtained from animals and human patients with sepsis and endocarditis, which indicates that E. faecalis of animal origin may be regarded as a hazard to human health. More studies are needed to quantify the animal reservoir in relation to E. faecalis infections in humans.
E. faecium and E. faecalis can survive in harsh environments, and good hygiene is therefore essential during food handling to minimize fecal contamination of the meat. Good hand hygiene and proper cleaning are also essential for patients to minimize the spread of enterococci in the clinical settings.
Footnotes
Acknowledgments
We thank James Suter, General Counsel at the Australian Pesticides and Veterinary Medicines Authority, for information on the use of avoparcin and virginiamycin in Australia, and Antonio Vieira, National Food Institut, The Technical University of Denmark, for information on the use of avoparcin in Latin America.
This work was part of The Danish Integrated Antimicrobial Resistance and Research Program (DANMAP) funded by the Danish Ministry of Health and Prevention and the European Union Sixth Framework Program “Approaches to Control Multiresistant Enterococci (ACE): Studies on Molecular Ecology, Horizontal Gene Transfer, Fitness and Prevention” under contract LSHE-CT-2007-037410.
Disclosure Statement
No competing financial interests exist.
Disclaimer
The information contained in this publication does not necessarily reflect the opinion or the position of the European Commission or the ECDC.