
Abstract
Vibrio parahaemolyticus, a common cause of foodborne gastroenteritis in people, is frequently isolated from a variety of seafood, including shrimp. The virulence of clinical V. parahaemolyticus strains is commonly associated with expression of thermostable direct hemolysin (TDH) and TDH-related hemolysin (TRH), which are encoded by the tdh and trh genes. This study was conducted to determine the prevalence rate of total and toxigenic V. parahaemolyticus in shrimp caught off the south coast of Iran. Three hundred freshly caught shrimp from four different species, Penaeus monodon, Penaeus semisulcatus, Penaeus indicus, and Penaeus merguiensis, were collected in three provinces along Persian Gulf in the south coast of Iran. Shrimp were collected at the end of each month from July 2008 to July 2009. The samples were analyzed for the presence of V. parahaemolyticus and the occurrence of toxigenic strains. Using conventional bacteriological techniques, 29 V. parahaemolyticus isolates were identified in which 28 strains were confirmed by a polymerase chain reaction assay targeting the tlh gene of V. parahaemolyticus. Using polymerase chain reaction assays targeting the tdh and trh genes, five (1.7%) and two (0.7%) isolates were tdh positive and trh positive, respectively. The tdh-positive isolates were commonly detected in summer, whereas no toxigenic strain was isolated in winter. To the best of our knowledge, the present study is the first report of the presence of toxigenic tdh- and trh-positive V. parahaemolyticus strains in the seafood in Iran.
Introduction
V
The bacterium generally inhabits coastal environments in tropical and temperate zones, and contaminates fishery products caught in these areas (Yano et al., 2006). V. parahaemolyticus is distributed throughout the world, but reported densities in the environment and in seafood vary greatly according to season, location, sample type, and analytical methodology employed (De Paola et al., 2003; Ristori et al., 2007).
The virulence of clinical V. parahaemolyticus strains is commonly associated with expression of thermostable direct hemolysin (TDH) (Nishibuchi et al., 1986), which is encoded by the tdh gene (Honda and Lida, 1993). V. parahaemolyticus can also produce a TDH-related hemolysin (TRH) encoded by the trh gene. Both tdh and trh genes are associated with illness due to V. parahaemolyticus infection. Although there is no biochemical method to detect TRH production, polymerase chain reaction (PCR) amplification can detect the presence of the trh gene. The gene for thermolabile hemolysin (tlh) has been shown to be present in all V. parahaemolyticus strains and is therefore used for the identification of the organism (Bej et al., 1999).
The epidemiology of vibriosis in Iran is essentially unknown, and to the best of our knowledge, the prevalence rate of toxigenic V. parahaemolyticus in shrimp in Iran has never been reported. The objective of the present study was to determine the prevalence rate of total and toxigenic V. parahaemolyticus in shrimp caught off the south coast of Iran.
Materials and Methods
Sample preparation
A total of 300 freshly caught shrimp of 4 different species, Penaeus monodon (72), Penaeus semisulcatus (80), Penaeus indicus (88), and Penaeus merguiensis (60), were collected from 3 provinces (Booshehr [130], Hormozgan [96], and Khoozestan [74]) along Persian Gulf in the south coast of Iran. Shrimp were collected at the end of each month from July 2008 to July 2009, placed in separate sterile plastic bags to prevent spilling and cross contamination, and immediately transported to the laboratory in a cooler with ice packs.
The samples were analyzed within 12 h of collection using aseptic techniques. First, the samples were placed in stomacher bags and homogenized at high speed for 90 sec. Then, 25 g of the homogenized sample was diluted with 225 mL of alkaline peptone water (Merck) containing 1% NaCl (pH 8.6) and incubated at 37°C for 6 to 8 h. After primary enrichment, thiosulfate citrate bile salt sucrose (TCBS) agar (Merck) was used as a selective medium for separation of Vibrio colonies. The samples were cultured at 37°C for 18 to 24 h using TCBS. The green or blue-green colonies (2 to 3 mm in diameter) grown on the TCBS agar plates were presumptively selected as V. parahaemolyticus colonies and were transferred to trypticase soy agar (TSA) plates (Merck) containing 3% NaCl. After incubation at 37°C for 24 h, the isolates were tested using conventional bacteriological methods, including Gram staining; culture in sulphide indol motility (SIM) and triple sugar iron agar (TSI) media; oxidase and catalase activity; growth in nutrient broth with 3%, 6%, 8%, and 10% NaCl; gelatin liquefaction (22°C); indol production; acid production from glucose, arabinose, mannitol, sorbitol, lactose and mannose, ornithine, arginine, and lysine decarboxylase; citrate utilization; and o-nitrophenyl-beta-D-galactosidase (ONPG) tests (Hosseini et al., 2004; Colakoglu et al., 2006).
Detection of V. parahaemolyticus from pure culture by PCR
One milliliter pure culture of V. parahaemolyticus, identified by the biochemical tests, was centrifuged at 13,000 g for 5 min at room temperature as previously described (Di Pinto et al., 2008). DNA purification was achieved using a Genomic DNA purification kit (Fermentas, GmbH) according to the manufacturer's instructions, and the total DNA was measured at 260 nm optical density according to the method described by Sambrook and Russell (2001).
The presence of tlh, tdh, and trh genes was detected according to the method described by Bej et al. (1999) and Tada et al. (1992). The PCR assays were performed using synthetic oligonucleotide primer pairs for tlh (5′-AAA GCG GAT TAT GCA GAA GCA CTG-3′ and 5′-GCT ACT TTC TAG CAT TTT CTC TGC-3′) (Bej et al., 1999), tdh (5′-GTA AAG GTC TCT GAC TTT TGG AC-3′ and 5′-TGG AAT AGA ACC TTC ATC TTC ACC-3′) (Tada et al., 1992), and trh (5′-TTG GCT TCG ATA TTT TCA GTA TCT-3′ and 5′-CAT AAC AAA CAT ATG CCC ATT TCC G-3′) (Tada et al., 1992). All oligonucleotide primers were obtained from a commercial source (CinnaGen). The reaction mixture for tlh analysis was performed in a total volume of 20 μL containing 1.2 μL of DNA sample, 11.9 μL of deionized water, 2 μL of 10 × buffer, 1.6 μL of 25 mM concentration of MgCl2, 1.6 μL of 2.5 mM concentration of dNTP, 0.8 μL (200 μM) of each primer, and 0.1 μL of 10 U/μL of Taq DNA polymerase (Roche Applied Science). The PCR assay was performed at 94°C for 3 min followed by 20 cycles of 94°C for 1 min, 60°C for 1 min, and 72°C for 2 min in a DNA thermal cycler (Master Cycler Gradient; Eppendrof ).
The DNA samples from all tlh-positive isolates were examined for tdh and trh genes with a multiplex amplification producing detectable amplicons at 250 and 373 bp, respectively. The PCR mixture for tdh and trh genes was prepared as described for the tlh gene. The PCR assay was performed at 94°C for 3 min and then for 35 cycles of 94°C for 1 min, 56°C for 1 min, and 72°C for 1 min.
Detection of V. parahaemolyticus from enrichment broth by PCR
One milliliter enrichment broth from each shrimp sample was centrifuged at 13,000 g for 5 min at room temperature. The cell pellets were subjected to DNA extraction as described above. Three milliliter aliquot of each sample was used for PCR amplification. All reactions were performed in triplicate.
The PCR-amplified products were examined by electrophoresis in a 1.5% agarose gel, stained with a 1% solution of ethidium bromide, and examined under ultraviolet illumination. A toxigenic V. parahaemolyticus strain possessing both tdh and trh genes and DNase-free water were used as the positive and negative controls, respectively.
Statistical analysis
Data were transferred to a Microsoft Excel spreadsheet (Microsoft Corp.) for analysis. Using SPSS 16.0 statistical software (SPSS Inc.), a chi-square test and a Fisher's exact two-tailed test analysis were performed, and differences were considered significant at values of p < 0.05.
Result
In the present study, the green or blue-green colonies on TCBS agar plates were detected in 118 of 300 (39.3%) shrimp samples. However, biochemical tests confirmed only 29 of 118 colonies as V. parahaemolyticus–positive samples. The molecular analysis carried out on these 29 strains gave positive results for 28 strains (Table 1) using a PCR assay targeting the tlh gene of V. parahaemolyticus (Fig. 1). The specificity of the PCR products was confirmed by sequence analysis (data not shown). In addition, five (1.7%) and two (0.7%) of V. parahaemolyticus isolates were tdh positive and trh positive, respectively, using PCR assays targeting the tdh and trh genes (Fig. 2). The PCR assays were performed in triplicate and no variability in the results was present. Overall, the observed difference in the frequency of detection of the tdh and trh genes from the pure culture versus the enrichment broth was not statistically significant. Also, the ratio of toxigenic to total V. parahaemolyticus isolates using the pure culture and the enrichment broth showed no statistically significant difference. In this study, no toxigenic V. parahaemolyticus was isolated from shrimp of P. merguiensis species.
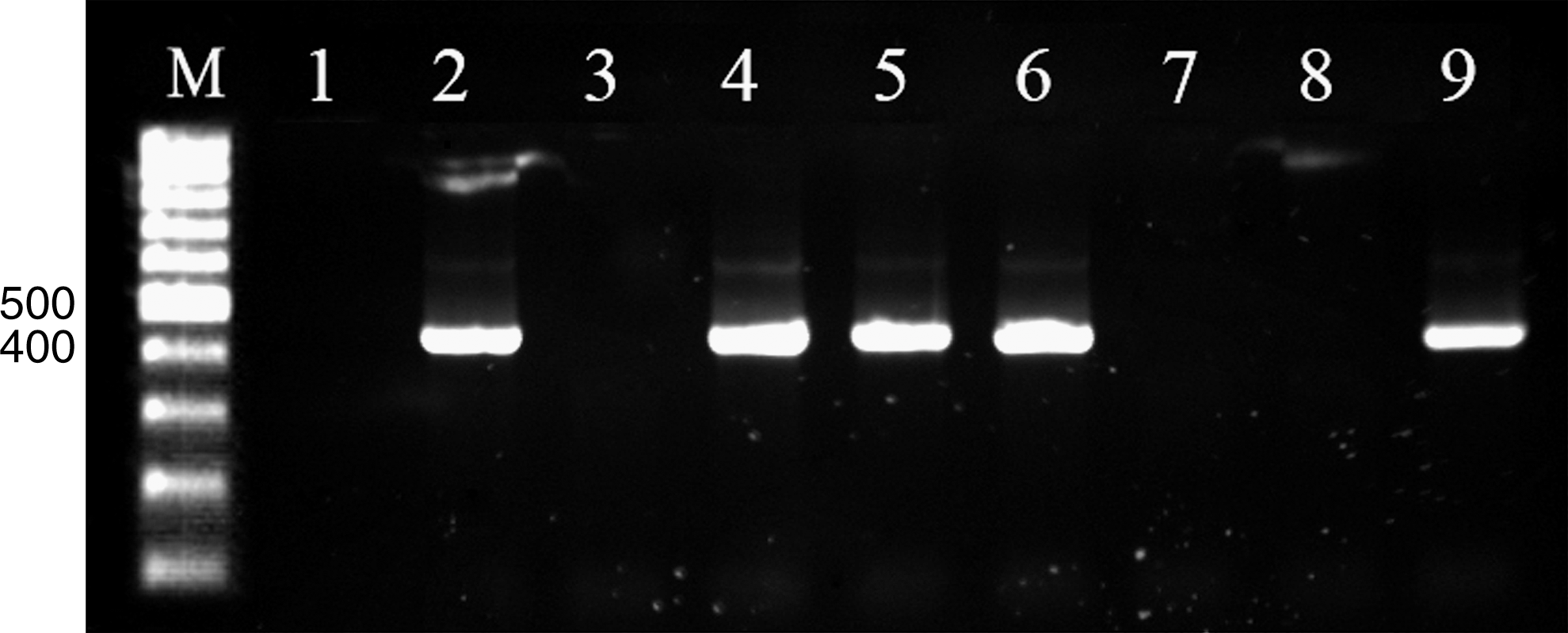
Ethidium-bromide-stained agarose gel polymerase chain reaction assay targeting the tlh gene. M: 100 bp DNA ladder; lane 1: negative control; lanes 2 and 4 to 6: positive samples; lanes 3, 7, and 8: negative samples; lane 9: positive control.
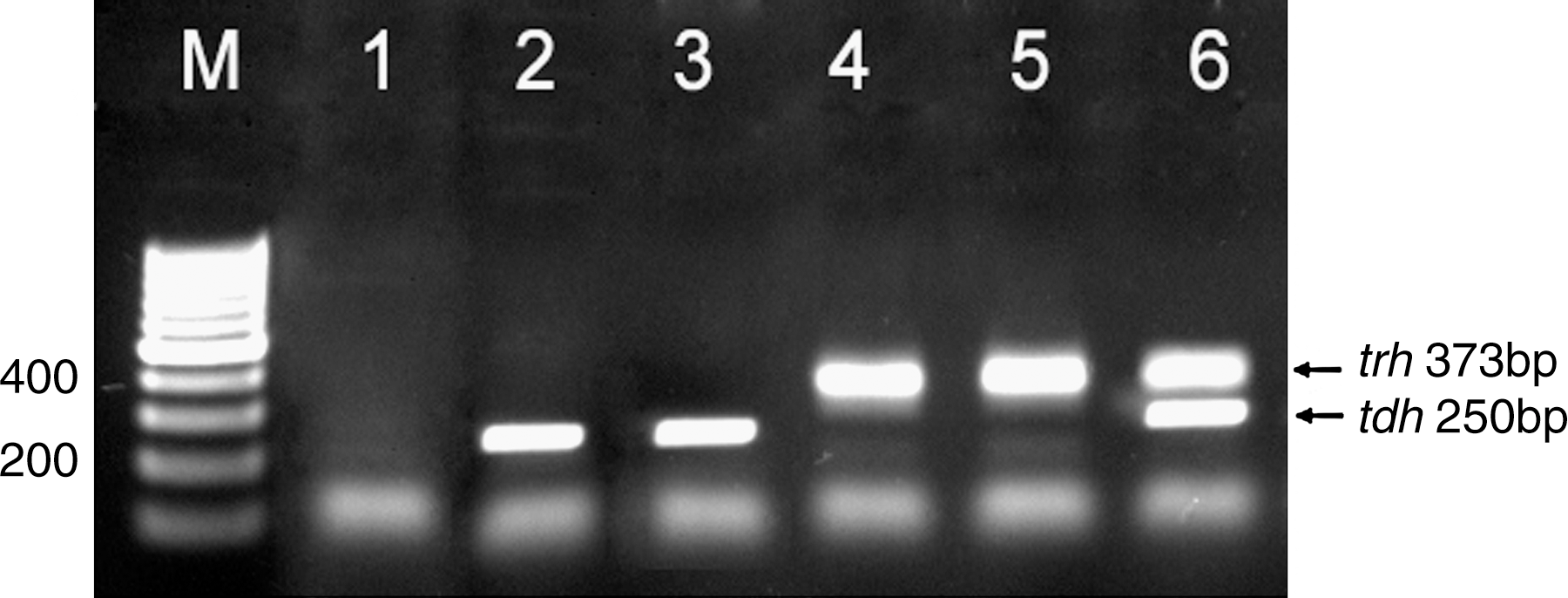
Ethidium-bromide-stained agarose gel polymerase chain reaction assays targeting the tdh and trh genes. M: 100 bp DNA ladder; lane 1: negative control; lanes 2 to 5: positive samples; lane 6: positive control.
The PCR assays performed on enrichment broth from each sample gave positive results for V. parahaemolyticus in 40 (13.3%) shrimp samples. TDH and TRH virulence factors were detected in seven (2.3%) and two (0.7%) samples, respectively.
The frequencies of total and toxigenic V. parahaemolyticus isolated from pure culture and enrichment broth at different provinces and seasons are shown in Tables 2 and 3, respectively. The prevalence rate of the tdh-positive isolates was higher in summer than in other seasons.
Discussion
Shrimp is one of the most important fishery products of Persian Gulf coastal provinces of Iran. Shrimp farming is an important industry of these provinces and a large portion of the products are export to other countries such as European Union countries (Hosseini et al., 2004). V. parahaemolyticus is one of the major causes of seafood-borne bacterial gastroenteritis and is frequently associated with the consumption of improperly cooked seafood (Deepanjali et al., 2005).
The prevalence rate of total V. parahaemolyticus of 9.3% (28/300) observed in shrimp in this study was in agreement with that recently reported in shrimp samples from India (Raghunath et al., 2008) and Malaysia (Sujeewa et al., 2009); however, it is lower than the results reported from Thailand (75.8%) (Wong et al., 1999), Taiwan (70.2%) (Wong et al., 1992), and China (39.4%) (Yang et al., 2008). Our results are higher than that reported from Turkey (0.8%) (Colakoglu et al., 2006) and from one previously reported from Iran (2.1%) (Hosseini et al., 2004).
In the present study the numbers of total and toxigenic V. parahaemolyticus isolates were higher during summer and fall, when the water temperature in Iran's southern coast is often higher than 25°C. However, in winter, when the water temperature drops to about 15°C, only three strains were isolated and none of them were found tdh/trh positive. The high frequency of V. parahaemolyticus isolation in shrimp samples during warmer months is in agreement with results reported in seafood by other investigators (Ottaviani et al., 2005; Mahmud et al., 2006; Di Pinto et al., 2008). However, no seasonal effect on the prevalence of V. parahaemolyticus in retail shrimp has been reported (Normanno et al., 2006).
The prevalence of tdh (5 of 28) and trh (2 of 28) carrying V. parahaemolyticus isolates reported in our study is comparable with those reported from Malaysia (Sujeewa et al., 2009). However, our results were higher than those of the study conducted in India in which 2 of 47 isolates were tdh positive and 1 of 47 isolates was trh positive (Gopal et al., 2005). In a study reported from China, 7.1% (2 to 28) of V. parahaemolyticus isolated from fresh shrimp were tdh positive, and no trh-positive sample was isolated (Yang et al., 2008). Raghunath et al. (2008) also reported that 25% of V. parahaemolyticus isolated from shrimp (Penaeus monodon) were trh positive, whereas no tdh-positive sample was found. In Italy, Di Pinto et al. (2008) also reported that 33.3% of V. parahaemolyticus isolated from seafood were tdh positive, and no trh-positive sample was reported. Up to 5% of the environmental Vibrio isolates may carry the tdh and/or the trh genes (Robert-Pillot et al., 2004). A frequency range of 0% to 12% of tdh-positive V. parahaemolyticus in environmental samples and seafood has also been reported (Ogawa et al., 1989; De Paola et al., 2003; Deepanjali et al., 2005). Variation in the prevalence of total and toxigenic V. parahaemolyticus isolates from raw shrimp samples reported in different studies may be a result of different sampling techniques, seasonal effects (Ottaviani et al., 2005; Mahmud et al., 2006; Di Pinto et al., 2008), and/or laboratory methodologies employed in different studies (bacteriological and biochemical testing vs. PCR assays). Further, a higher prevalence rate of V. parahaemolyticus–positive in seafood could be due to cross contamination during manual processing or insufficient hygiene during storage and transport in the seafood markets.
The results of this study indicate that the conventional bacteriological techniques based on identifying phenotypic characteristics are often unable to identify V. parahaemolyticus. In addition, in comparison to molecular analysis such as PCR targeting the tlh gene, the conventional methods for detecting V. parahaemolyticus are technically time consuming and labor intensive. Detection of V. parahaemolyticus by PCR applied directly to enrichment broth has been shown to be highly specific and may help to avoid any interference with competitive Vibrio alginolyticus colonies (Di Pinto et al., 2008).
V. parahaemolyticus is inactivated by most heat treatments applied to seafoods. Since cross contamination of other foods due to contact with contaminated seafood is possible, prevention of cross contamination from raw seafood to cooked product is important. It is also recommended to process and storage raw seafood at low temperature to prevent growth of V. parahaemolyticus.
To the best of our knowledge, the present study is the first to report the presence of toxigenic tdh- and trh-positive V. parahaemolyticus strains in the seafood in Iran. Considering the widespread distribution in marine environments, fast replication times, and low infectious doses of the pathogenic strains (Kaysner and DePaola, 2001), intensive and continuous monitoring of potentially pathogenic V. parahaemolyticus is strongly recommended to assess the human health risk arising from seafood consumption. The results of this study emphasize the need for an efficient survey of the total and toxigenic V. parahaemolyticus present in seafood in Iran. Further, the food safety problem associated with V. parahaemolyticus in seafood necessitates developing specific risk management guidance information for this pathogen and governmental guidelines for vibrios in postprocessing or ready-to-eat seafoods in Iran.
Footnotes
Acknowledgments
The authors would like to thank Mahdi Raisi, Hasan Mommtaz, and Manouchehr Momeni for the sincere help in performing technical parts of the project.
Disclosure Statement
No competing financial interests exist.