
Abstract
The objective of this cross-sectional study was to compare the prevalence of Salmonella and antimicrobial-resistant Salmonella, as well as investigate the distribution of this pathogen in organic and conventional broiler poultry farms. Fecal (n = 420), feed (n = 140), and drinking water (n = 140) samples were collected from birds at 3 and 8 weeks of age for 2-flock cycles. One house was sampled per farm at three organic and four conventional broiler farms from the same company in North Carolina. All samples were analyzed for the presence of Salmonella using selective enrichment techniques. Further phenotypic (antimicrobial susceptibility) and genotypic (pulsed-field gel electrophoresis [PFGE]) testing were performed. Salmonella prevalences in fecal samples were 5.6% (10/180) and 38.8% (93/240) from organic and conventional farms, respectively. From feed, 5.0% (3/60) and 27.5% (22/80) of the samples were positive for Salmonella from organic and conventional farms, respectively. None of the water samples were positive for Salmonella. Seventy isolates were characterized by antimicrobial susceptibility and PFGE types. The two most common resistance phenotypes were single resistance to streptomycin (36.2% [25/58]: conventional; 25% [3/12] organic), and multidrug resistance to six antimicrobial agents: ampicillin-streptomycin-amoxicillin/clavulanic acid-cephalothin-ceftiofur-cefoxitin (AmStAxChCfFx; 39.7%: conventional only). Genotypic analysis using PFGE showed clonality among isolates within and between the two types of farms. The results of our study suggest that within this poultry company, the prevalence of fecal Salmonella was lower in certified-organic birds than in conventionally raised birds, and the prevalence of antimicrobial-resistant Salmonella was also higher in conventionally raised birds than in certified-organic birds.
Introduction
A
Several studies have assessed preharvest Salmonella prevalence on conventional broiler farms (Renwick et al., 1992; Bailey et al., 2001; Liljebjelke et al., 2005; Rodriguez et al., 2006; Arsenault et al., 2007). However, very little is known about the prevalence of Salmonella on large-scale USDA-certified organic broiler farms. In a recent study by Siemon et al. (2007), the prevalence of fecal Salmonella in pasture chicken farms (16%) was lower than the prevalence in conventional chicken farms (30.0%). Pasture broiler farms are small operations (average of 500 birds raised per year) where chickens are reared outside on pasture in open-air moveable pens (Jacob et al., 2008). Pasture birds are grown without subtherapeutic (growth promotion) or therapeutic antimicrobial use (i.e., considered as antibiotic-free chickens); however, they are fed commercial nonorganic diet. The USDA has placed a set of national standards the broiler production must meet to be labeled organic (Dimitri and Greene, 2002). These standards are primarily (1) birds must be raised without the use of antibiotics, (2) fed organic dietary supplements and consume all organic feed free of animal by products, and (3) have access to the outside environment (Dimitri and Greene, 2002). Organic poultry production focuses on animal health and welfare, good management practices, and product quality, whereas, conventional poultry production focuses on reducing costs and maximizing production through weight gain and feed efficiency (Sundrum, 2006).
To the best of our knowledge, this is the first study conducted to compare Salmonella prevalences, antimicrobial-resistant phenotypes, and dissemination in large-scale USDA-certified organic broiler chicken farms compared to conventional broiler farms.
Materials and Methods
Study design and sample collection
One poultry company in North Carolina that operates both USDA-certified organic and conventional broiler farms participated in this study. On organic broiler farms, birds were reared in medium-sized houses (∼200–300 feet long) that are similar in structure to conventional farms with an all-in–all-out system, fed all USDA-certified organic feed free of antimicrobial agents and animal by-products, exposed to sun-light, have twice more area per square foot than conventional birds, and have access to the outside environment, although birds usually prefer to stay in-doors. Broiler chicks were brought to the farms at 1 day of age and were sent to slaughter at 55–60 days of age. Houses had from 3000 to 5000 birds inside, per flock. On conventional poultry farms, the broilers were reared intensively in large houses (∼500–600 feet long) with an all-in–all-out system. Houses had from 15,000 to 30,000 broilers inside, and the broilers were slaughtered at 50–55 days of age. Feed-grade antibiotic (i.e., bacitracin methylene disalicylate) was routinely used at 25–50 g/ton of feed on the conventional poultry farms. Further, coccidiostats were used on the conventional farms.
A convenience sample of seven broiler farms (three organic and four conventional) were included in the study. One broiler house per farm (organic and conventional) was sampled. Each farm/house included in the study was followed for two consecutive flock cycles. Each broiler flock within a house was sampled twice; at 3–4 weeks of age and 1 week before slaughter to approximate Salmonella prevalence near slaughter. Appropriate dress in clean coveralls, plastic shoe covers, and lab-grade plastic gloves was used on entry to the farm. At each visit, 15 individual fresh fecal floor droppings (∼5 g) were collected using a zig-zag pattern through the entire house at the organic and conventional farms. In addition, we collected feed samples (n = 5; 50–100 g) and drinking water (n = 5; 1200 mL). Feed samples consisted of feed hopper (n = 2) and feed lines (n = 3). A Feed hopper is a V-shaped container for the incoming feed to the house to provide feed to the birds through the feed lines. Feed was collected as soon as new feed is dispensed into the hopper. Water samples consisted of house main water line (n = 1) and in-house drinking nipples (n = 4). Fecal and feed samples were collected with the use of sterile plastic gloves and sealed in sterile Whirl Pak© bags (Wisconsin; Nasco, Ft. Atkinson, WI), and water samples were collected in sterile Nalgene® containers (Nalgene, Rochester, NY). All samples were put in coolers on ice and shipped to the University of Georgia Center for Food Safety for laboratory analysis within 36–48 h of collection.
Salmonella isolation and identification
Fecal samples were mixed thoroughly and 1 g portions were added to 9 mL tetrathionate brilliant green broth (TBG) with 2 mL iodine (Difco, Division of Becton, Dickinson and Co., Sparks, MD) and incubated for 24 h at 42°C. Two loopful of the incubated media were streaked onto two xylose-lysine-tergitol-4 agar plates (Difco) for selective differentiation via 24 h incubation at 37°C. One presumptive black colony from each positive xylose-lysine-tergitol-4 plate was tested for biochemical reaction on triple sugar iron (TSI; Difco) slant. The slants were incubated at 37°C for 24 h. Identification of suspected Salmonella was confirmed and serogrouped using poly-O Salmonella-specific antiserum (MiraVista, Indianapolis, IN). A portion of the growth from the TSI slants was transferred and streaked into nutrient agar slants (Difco) and incubated at 37°C for 24 h. Nutrient agar slants with Salmonella growth were stored at room temperature until shipped to The Ohio State University Infectious Disease Molecular Epidemiology Laboratory for antimicrobial susceptibility testing and pulsed-field gel electrophoresis (PFGE) genotypic characterization.
A 25 g portion of feed sample was mixed with 225 mL TBG with 4 mL iodine (Difco), and incubated for 24 h at 42°C. The remaining of the isolation procedure was the same as the method used for isolation from fecal samples. Water samples were cultured using two methods: (1) 100 mL aliquot of water sample was added to 200 mL TBG with 4 mL iodine, and incubated for 24 h at 42°C, and (2) using Moore swab method (Moore, 1948), where a sterile cotton swab (four-inches-by-three-feet-long gauze) was suspended in the 1000 mL of water sample with stirring and left for 24 h at room temperature (24°C), and then swab was aseptically removed, cut, and added to 225 mL TBG with 4 mL iodine, and then incubated for 24 h at 42°C. The remainder of the isolation procedure for both methods was the same as for isolation from fecal samples. Salmonella isolates (n = 70) were serogrouped using poly-O Salmonella-specific antiserum (MiraVista).
Antimicrobial susceptibility test
Antimicrobial susceptibility testing of Salmonella isolates (n = 70) was performed using Kirby-Bauer disk diffusion method to a panel of 16 antimicrobial agents all of which are routinely tested by the National Antimicrobial Resistance Monitoring System (CDC, 2003). The following BBL™ Sensi-Disc™ antimicrobial susceptibility test discs (Becton, Dickinson) with their respective disc potencies were used: ampicillin (Am-10 μg), amoxicillin/clavulanic acid (Ax-20/10 μg), amikacin (An-30 μg), cefoxitin (Fx-30 μg), ceftriaxone (Ce-30 μg), ceftiofur (Cf-30 μg), cephalothin (Ch-30 μg), chloramphenicol (Cl-30 μg), ciprofloxacin (Cip-5 μg), gentamicin (Gm-10 μg), kanamycin (Km-30 μg), nalidixic acid (Nl-30 μg), streptomycin (St-10 μg), sulfamethoxazole (Su-250 μg), tetracycline (Te-30 μg), and Trimethoprim/sulfamethoxazole (Sm-1.25/23.75). Results were interpreted according to the Clinical Laboratory Standards Institute (CLSI) recommendations (NCCLS, 2002). All isolates that showed intermediate resistance were grouped with the susceptible strains to avoid overestimation of resistance. Control tests of E. coli ATCC 25922, E. coli ATCC 35218, Enterococcus faecalis ATCC 29212, Staphylococcus aureus ATCC 25923, and Pseudomonas aeroginosa ATCC 27853 were regularly performed in accordance with the CLSI standards (NCCLS, 2002).
Genotypic characterization of Salmonella using PFGE
DNA fingerprinting of Salmonella isolates using PFGE were performed according to CDC PulseNet standardized protocol (Ribot et al., 2006). Briefly, an overnight grown Salmonella cell concentration was adjusted by diluting with sterile cell suspension buffer to the OD value 1.3 to 1.4 measured at 610 nm wavelength with a spectrophotometer. Agarose-embedded cells were lysed, and intact genomic DNA was digested with 50U of XbaI restriction enzyme (New England Biolabs, Ipswich, MA) for 2 h at 37°C. The fragments were then separated by CHEF-DR® III system (Bio-Rad Laboratories, Hercules, CA) with the following conditions and reagents: 1% SeaKem Gold agarose (FMC BioProducts, Rockland, ME), in 0.5% Tris-borate EDTA buffer; temperature, 14°C; voltage, 6 V/cm; run time, 18 h with switch times ranging from 2.2 to 63.8 s. The PulseNet “universal” standard marker strain Salmonella enterica serovar Braenderup H9812 was used as a molecular reference marker and an “Out-group” strain. The gels were stained with ethidium bromide, and the DNA bands were observed under UV trans-illumination (Gel Doc™ 2000; Bio-Rad Laboratories) and gel images were captured using Quantity one 1-D analysis software (Bio-Rad Laboratories).
Fingerprint images were analyzed by Bionumerics software V. 4.61 (Applied Maths, Kortrijk, Belgium) as per the manufacturer's recommendation. Briefly, the captured gel images were normalized by aligning the lanes of the gel to a reference standard for the Salmonella database. To identify both the distance measure and cluster algorithm that “best” describe clustering among PFGE gel patterns, various distance (i.e., proximity) measures (e.g., Dice, Jaccard, and Matching) and clustering algorithms (e.g., complete linkage, single linkage, and Ward) were applied, and a dendrogram tree was generated. A dice coefficient index with optimization of 1.5% and position tolerance of 2.0% was used in analysis of images in this study. For clustering purposes, a threshold cut-off value of 80% was used with subclustering at 85%.
Statistical analysis
The association between fecal Salmonella prevalence and the study factors (farm type [organic and conventional], age group [3–4 weeks, and 1 week before slaughter], and flock cycle [first and second]) in the present study was assessed using a generalized linear model, with binomial error distribution and logit link function and adjusted for dependency within farms, using a generalized estimated equations (GEE) in STATA software version 10.1 (Stata Corp., College Station, TX). GEE is a multivariable logistic regression with a population-averaged model that adjusted for estimated correlations among the isolates in this study within a cluster (i.e., farms). The reported odds ratios (OR) from GEE was comparing the odds of Salmonella in a group, while adjusted for the dependence of isolates within farms, to the odds of Salmonella in the other group (Dohoo et al., 2003).
The 16 antimicrobial-resistant Salmonella outcomes (binary), as well as the multidrug resistance totals (multinomial), were cross-tabulated with farm type. The proportion of Salmonella isolates resistant to each of the antimicrobial agents was compared by farms type using either a two-sided 2-by-2 Fisher's exact test or 2-by-n likelihood ratio chi-square test, as appropriate, with STATA software. Multidrug resistance (resistance ≥2 agents) was assessed by farm type across all isolates, using an m × n likelihood ratio chi-square test.
Results
Salmonella prevalence
A total of 700 samples (300 organic and 400 conventional) were collected from three organic and four conventional broiler farms within one poultry company over two consecutive flock cycles. The overall prevalence of Salmonella across all farms, sample types, and age group was 4.3% (13/300) in organic broiler farms compared to 28.8% (115/400) in conventional broiler farms. Table 1 shows Salmonella prevalence by sample type for each of the seven farms that participated in the study. The prevalence of Salmonella by sample type was compared between organic and conventional broiler farms as shown in Table 2. The OR for prevalence of Salmonella in fecal and feed samples adjusted for the dependence of isolates within farms was significantly (p < 0.05) higher (OR = 11.9 and 7.2, respectively) in conventional farms than in organic farms. Salmonella isolates from organic feed were all from the feed lines (i.e., feed pans); however, two of the isolates from conventional feed were from the feed hopper (i.e., incoming feed). The prevalence of Salmonella by birds' age group and by flock cycle was compared between organic and conventional broiler farms as shown in Table 3. In Table 3, the OR for prevalence of Salmonella in samples collected from birds 1 week before slaughter adjusted for the dependence of isolates within farms was significantly (p < 0.05) higher (OR = 2.2) than 3–4 weeks of age in conventional farms, whereas no significant difference (p > 0.05) in organic farms was found. Further, within each farm type, the OR for prevalence of Salmonella in samples collected from the second flock cycle, adjusted for the dependence of isolates within farms, was not significantly (p > 0.05) different compared to first flock cycle.
CK, KB, PG, JDK, MH, RS, and RG are codes used in the study for farm identification during sample collection and analysis.
Frequencies are contrasted by each farm (organic and conventional) and sample type across all age groups and flock cycles.
Odds ratio values represent a comparison of the odds of the prevalence of Salmonella in fecal and feed samples, adjusted for the dependence of isolates within farms, in conventional compared to organic farms.
p-Values are adjusted for the dependence of Salmonella isolates within farms by using generalized estimating equation statistic in STATA (Stata Corp.). Frequencies are contrasted by sample type across all farms, age group, and flock cycles.
Odds ratio values represent a comparison of the odds of the prevalence for each age group within organic and conventional Salmonella isolates.
p-Values are adjusted for the dependence of Salmonella isolates within farms by using generalized estimating equation statistic in STATA (Stata corp.). Frequencies are contrasted by age group across all farms, sample type, and flock cycles.
Odds ratio values represent a comparison of the odds of the prevalence of Salmonella, within each farm type, in samples collected from the second flock cycle adjusted for the dependence of isolates within farms in conventional compared to organic farms.
The majority of the isolates (92.9%; 65/70) were Salmonella serogroup C. The antisera used in the laboratory did not distinguish between C1, C2, or C3, so we only know that it was group C. The remaining five isolates were serogroup B.
Antimicrobial resistance
Antimicrobial susceptibility testing was performed on 70 selected Salmonella isolates (12 from organic and 58 from conventional farms) out of 128 isolates that represented the majority from both types of farms. These isolates were selected to represent all the farms, farm type, and sample type (if available). The individual antimicrobial resistance Salmonella phenotypes cross-tabulated by farm type are shown in Table 4.
p-Values are based on Fisher's exact test of the differences in risk between farm type.
In isolates from organic farms, 25% (3/12) were pansusceptible, 33.3% (4/12) had single-agent-resistance, and 41% (5/12) were resistant to two or more antimicrobial agents. In conventional farms, 1.7% (1/58) of the isolates were pansusceptible, 36.2% (21/58) were single-agent-resistant, and 62% (36/58) were resistant to two or more antimicrobial agents. The distribution of multidrug-resistant Salmonella phenotypes by farm type is shown in Table 5. The proportion of multidrug-resistant Salmonella isolates differed significantly by farm type (p = 0.001). The predominant resistance patterns among Salmonella isolates from conventional farms were ampicillin- streptomycin-amoxicillin/clavulanic acid-cephalothin-ceftiofur-cefoxitin (AmStAxChCfFx; 39.7%: 25/58), and streptomycin (36.2%; 21/58), whereas the predominant Salmonella resistance patterns from organic farms were pansusceptible (25%; 3/12), and streptomycin (25%; 3/12).
Am, ampicillin; Ax, amoxicillin/clavulanic acid; An, amikacin; Fx, cefoxitin; Ce, ceftriaxone; Cf, ceftiofur; Ch, cephalothin; Cl, chloramphenicol; Cip, ciprofloxacin; Gm, gentamicin; Km, kanamycin; NI, nalidixic acid; St, streptomycin; Su, sulfasoxazole; Te, tetracycline, Sm, Trimethoprim/sulfamethoxazole.
Pulsed-field gel electrophoresis
To assess genotypic relatedness, PFGE was used to analyze 70 Salmonella isolates. A total of four main PFGE clusters were found in this study (Fig. 1a, b). Cluster type 1 (with two subclusters 1A and 1B) was the most predominant. This cluster was composed of isolates that originated from both production types and various resistance phenotypes. The PFGE patterns showed the highly clonal nature of the Salmonella isolates within and between the organic and conventional broiler farms. In addition, clonality of isolates between feed and fecal samples was detected at individual farm levels implying potential transmission of Salmonella via contaminated feed.
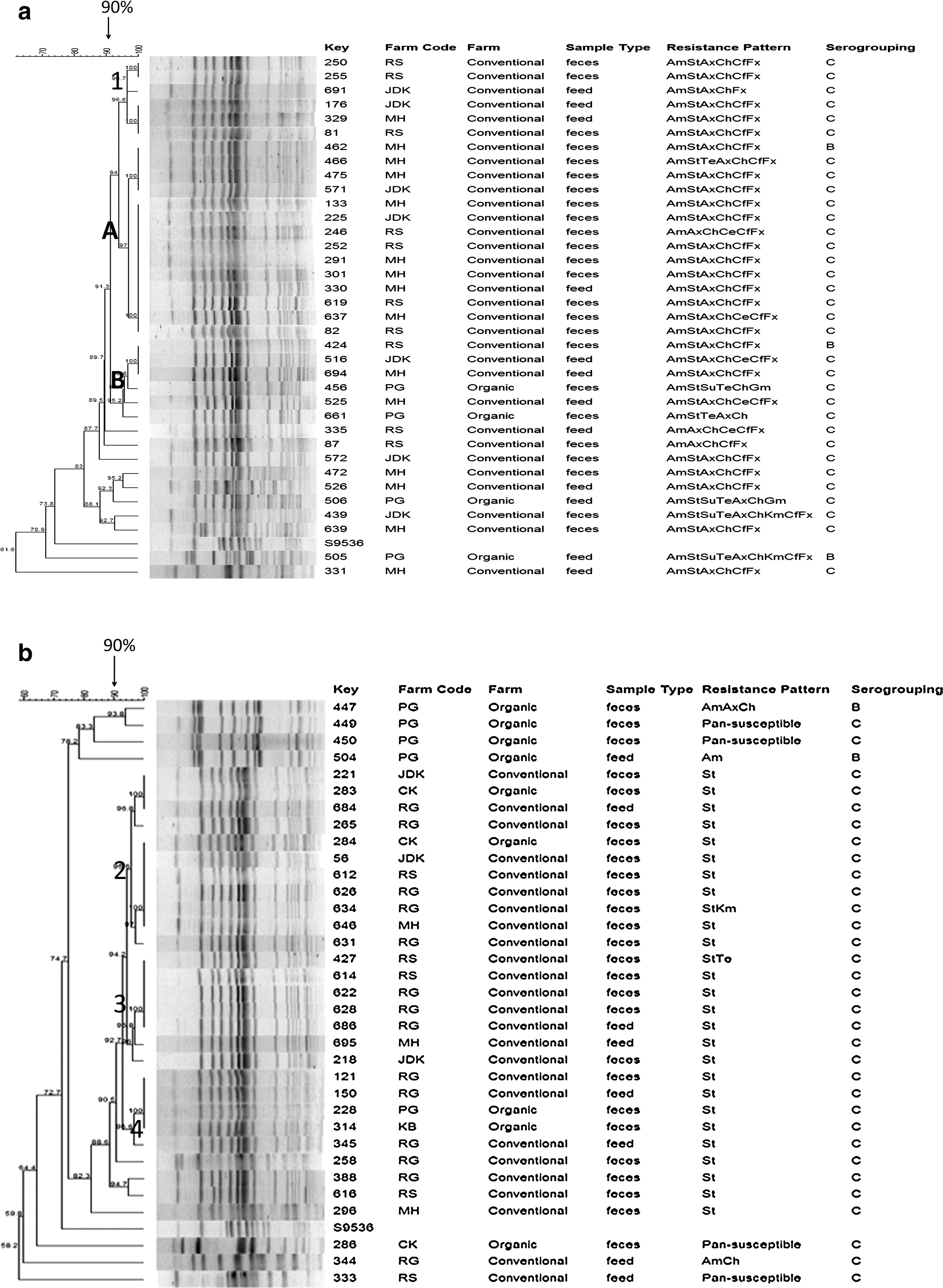
Dendogram of pulsed-field gel electrophoresis patterns of Salmonella isolates (n = 70) recovered from feces and feed sampled from organic and conventional broiler farms. (
Discussion
To the best of our knowledge, this present study is the first to investigate the prevalence of Salmonella, determine antimicrobial-resistant phenotypes of Salmonella, and investigate Salmonella dissemination in large-scale USDA-certified organic broiler farms compared to conventional broiler farms owned by the same company.
In the current study, the overall prevalence of Salmonella and fecal shedding of Salmonella was lower in organic farms than in conventional farms. Further, the overall prevalence of Salmonella by age group (3–4 weeks and 1 week before slaughter) was significantly different within conventional farms, but not significant in organics. There was no significance difference in the prevalence of Salmonella by flock cycle for either farm type. In feed, the prevalence of contamination with Salmonella was lower in organic farms than in conventional farms. Contaminated feed at the organic farms was only present in the feed lines (i.e., feed pans), which is likely due to feed contamination with the birds' fecal droppings. Salmonella was present in feed samples from the feed hopper (i.e., incoming feed) as well as in the feed line samples at the conventional farms. This may indicate that incoming conventional feed was contaminated with Salmonella. Some Salmonella cells could be injured during feed production (e.g., heating step during feed pelleting) of conventional feed. Organic feed was produced without pelleting (i.e., mash). We used a selective enrichment broth (TBG) to recover Salmonella from both types of feed samples. This method might underestimate the true prevalence of cultivable Salmonella, mostly in conventional feed samples. Despite the use of selective broth, we recovered higher proportions of Salmonella from conventional feed than from organic feed samples.
The presence of Salmonella in broiler feed and feed ingredients (e.g., bone, meat, and fish meal) is well documented (Allred et al., 1967; Stott et al., 1975; Furuta et al., 1980; Bailey et al., 2001; Maciorowski et al., 2004). Bailey et al. (2001) reported Salmonella prevalence of 2.33% (6/258) and 2.28% (6/263) in feed hopper and feeder (i.e., feedlines) in conventional broiler farms. In our study, it was common to find fecal droppings in the feed pans at both types of farms; however, due to the higher prevalence of fecal shedding of Salmonella at the conventional farms (as our results indicated), conventional feed was more likely to be contaminated with fecal Salmonella and horizontally spread this pathogen to other birds within the same house. In a previous study by Siemon et al. (2007), the authors reported that fecal Salmonella prevalence in conventional bird flocks (30%; 125/419) was significantly higher than in pasture flocks (16%; 83/512). The conventional and pasture farms were owned by different companies. Pasture poultry farms are small operations where birds are reared on pasture and usually not classified as USDA-certified organic birds (Jacob et al., 2008). Various overall estimates of Salmonella prevalence in conventional broiler farms in the United States were reported in several studies ranged from 10% to 26% (Bailey et al., 2001; Liljebjelke et al., 2005; Rodriguez et al., 2006). These prevalence estimates usually vary between studies due to seasonal effects, differences in the hatchery sources, feed composition, vaccination programs, and flock-disease status. It has been shown that Salmonella is found in the hatcheries due to vertical and/or horizontal transmission of this pathogen (Cox et al., 1990; Bailey et al., 1992; Bailey et al., 1994; Cox et al., 1995). Broiler chicks (organic or conventional) in this study were originated from the same conventional hatchery, but most likely from different breeder flocks. The cooperator poultry company has no information on which broiler birds came from which breeder flocks (personal communication with the poultry company's consultant veterinarian). There was no sample collection conducted at the arrival of chicks to the farms, as the main interest of the study was to determine Salmonella prevalence, Salmonella antimicrobial resistance profiles, and dissemination in the middle of the production cycle as well as during the last week before slaughter to assess the preharvest food safety risk associated with organic broilers compared with conventional broilers. Organic birds were raised under organic conditions starting at day 1 when they arrived at the farms. If we assume that the distribution of chicks at both farm types was random, then each farm (organic or conventional) would have received broiler birds representing the multiple breeder flocks that supplied the hatchery.
The prevalence of Campylobacter in large-scale USDA-certified organic and conventional farms in Ohio was estimated at 89% and 66%, respectively (Luangtongkum et al., 2006). The authors collected their study samples from intestinal tracts of organic and conventional birds at processing plants. In a different species, prevalence of Salmonella was higher in antibiotic-free (niche-market and out-door) swine farms than in conventional (intensive and indoor) swine farms (Gebreyes et al., 2006; Gebreyes et al., 2008).
We did not detect Salmonella in our water samples using either isolation method. In Lilebjelke et al. (2005), the authors found one Salmonella isolate out of 56 water samples collected from conventional broiler houses. They used a similar enrichment/isolation protocol (100 mL of water added to 200 mL of TBG with 4 mL iodine) as in our study. In another study, Bailey et al. (2001) reported a Salmonella prevalence of 1.4% (10/731) in water sampled from house waterlines. We believe that if Salmonella was present in the water samples, it would have been at very low levels below the detection limit (10 CFU/mL) of the two isolation/culture methods used in the present study.
A comparison between Salmonella prevalence in organic and conventional broiler meat at retail stores was examined in several studies. Lestari et al. (2009) reported Salmonella prevalence in organic (20.8%) and conventional (22%) chicken carcass samples collected from 27 retail stores in Baton Rouge, Louisiana. The authors found that the difference was not statistically significant between both types of samples. It was found in other studies that Salmonella prevalence in broiler chicken meat from conventional birds was lower than that in chicken meat from organic birds (Bailey and Cosby, 2005; Cui et al., 2005). Cui et al. (2005) reported Salmonella prevalence in organic and conventional chicken meat, collected from retail stores in Maryland, at 61% and 44%, respectively. It was not clear in the two studies (Cui et al., 2005; Lestari et al., 2009) whether organic chicken carcasses were from USDA-certified organic birds, pasture birds, or free-range birds. In Bailey and Cosby (2005), the authors reported an overall Salmonella prevalence of 31% and 25% in free-range and all-natural chicken meat, respectively, obtained from retail stores. Further, it was reported that the prevalence of Salmonella in their USDA-certified free-range chicken samples was 60%. In that study, the prevalence of Salmonella estimates at 31% and 25% were compared to the Food Safety and Inspection Service Salmonella reports during chicken processing (USDA, 2004). The Salmonella prevalence was 12.8% in conventional chicken carcasses according to the Food Safety and Inspection Service estimates in 2004, and most recently the postchill prevalence of Salmonella was reported at 5.2% (n = 2114) (USDA, 2009).
The overall prevalence of individual and multidrug antimicrobial resistance was higher in Salmonella isolates from conventional broiler farms than in those from organic broiler farms. Multidrug resistance (resistance to two or more antibiotics) was more frequent in Salmonella isolates from conventional broiler farms (55.2%) compared with organic farms (41.6%). Forty-three percent (25/58) of isolates from conventional farms were resistance to six drugs (AmStAxChCfFx). The prevalence of multidrug-resistant Salmonella at conventional and organic farms in the study of Siemon et al. (2007) was only 35% and 30.2%, respectively. Conventional farms in the present study used bacitracin methylene disalicylate antibiotic supplement in feed at a subtherapeutic level to prevent necrotic enteritis infections caused by Clostridium perfringens and as a growth promoter. In contrast, organic farms did not use any antibiotics as a supplement or as a treatment. Necrotic enteritis is a multifactorial disease with a complex etiology. Although it would be interesting to compare the prevalence of Clostridium perfringens and clinical evidence of necrotic enteritis in both farm types, we did not assess that as it was beyond the scope of our study. The predominant resistance patterns in Salmonella isolates from conventional farms were AmStAxChCfFx and single-resistance to streptomycin. The predominant multidrug resistance pattern in Siemon et al. (2007) in conventional farms was AmStAxCSuTe (C, chloramphenicol; Su, sulfasoxazole; Te, tetracycline), which was similar to ours except for the cephalosporins class resistance. Many of the multidrug-resistant Salmonella isolates in our study have unique cephalosporins drug-class resistance pattern (cephalothin, cefoxitin, ceftiofur, and ceftriaxone). From a public health standpoint, ceftriaxone is considered to be a drug of choice to treat human cases of salmonellosis, especially in children. Ceftriaxone resistance in Salmonella in chicken is not common as it was found in 1% isolates out of 1121 (Gray et al., 2004). In our study, resistance to ceftriaxone was found in 8.6% (5/58) of Salmonella isolates, all were from conventional farms. Cui et al. (2005) found that 3.3% and 54% of Salmonella isolates from organic and conventional retail chicken were resistant to cephalothin-cefoxitin-ceftiofur, respectively. Similar findings on decreased susceptibility to cephalosporins in Salmonella isolates from retail organic and conventional chickens were reported by Lestari et al. (2009). Single resistance to streptomycin was higher (91.4%; 53/58) among isolates from conventional farms in the current study than 0.5% in Siemon et al. (2007) conventional farm samples. In our organic farms, the predominant Salmonella resistance patterns were pansusceptible (25%; 3/12) and streptomycin (25%; 3/12). Siemon et al. (2007) reported higher percentage of pansusceptible Salmonella isolates from pasture broilers (57%) and lower for streptomycin (3.1%) compared to our findings in organic broilers. The prevalence of individual and multidrug-resistant Salmonella from our organic farms need to be interpreted with caution as the number of the isolates (n = 12) from the collected samples (n = 300) was low. However, this suggested that the preharvest risk of Salmonella shedding in organic broiler farms was much lower than in the conventional broiler farms. According to the recently released NARMS (2006) report, Salmonella prevalence in retail chicken breast was 12.7% with 2.0% resistant to ACSSuTAuCf (ampicillin, chloramphenicol, streptomycin, sulfamethoxazole/sulfisoxazole, tetracycline, amoxicillin-clavulanic acid, and ceftiofur).
The PFGE genotyping findings suggested a highly clonal pattern for Salmonella isolated from organic and conventional broiler farms. Salmonella dissemination was not only within individual farms, but also between farms despite the differences in antimicrobial resistance patterns between the isolates. The resistance phenotypes in some or most cases may be inducible via antimicrobial use. Designated poultry company personnel were allowed to work with multiple farms (both organic and conventional) to supervise broiler production and assist growers. Further, farms sampled were located within a 30-mile radius. The high clonality between isolates of fecal and feed origin is significant as it may imply one additional route of dissemination of Salmonella strains to multiple farms via contaminated feed. Another possible explanation for the high clonality among the isolates is the lack of diversity among Salmonellae in our study. Around 93% of our isolates were Salmonella group C. Further, according to the literature, the majority (80%–90%) of Salmonella isolated from broiler birds are Salmonella Kentucky, which belongs to group C. However, due to the budget limitations for this study, we did not serotype any of the isolates. In another study, Lestari et al. (2009) found a very similar PFGE pattern in Salmonella isolates from organic and conventional chicken meat collected at retail stores.
Conclusions
Our study aimed to compare the prevalence of Salmonella in large-scale USDA-certified organic broiler farms relative to conventional broiler farms, within the same poultry company in North Carolina. The results of our study suggest that within this poultry company, prevalence of fecal Salmonella was lower in certified-organic birds than in conventionally raised birds. In addition, the prevalence of antimicrobial-resistant Salmonella was higher in conventionally raised birds than in certified-organic birds. Multidrug-resistant Salmonella strains were isolated from organic broiler farms even with the absence of antimicrobial selection pressure. Similarities in PFGE patterns for Salmonella isolates from both farm types may suggest that these organisms were circulating within the poultry company's farms or due to the lack of diversity among the Salmonella isolates in our study. Further longitudinal studies are needed to determine Salmonella loads quantitatively and verify Salmonella serotypes in multiple large-scale certified-organic on-farm operations and at processing plants.
Footnotes
Acknowledgments
We thank the administrators of the poultry company in North Carolina for their cooperation in conducting this research. We also thank Rebekah Turk, Kim Hortz, Aparna Petkar, and David Mann for their technical assistant in processing samples and performing microbiological assays at our laboratory. We also thank Dr. Wondwossen Gebreyes of The Ohio State University and his Infectious Disease Molecular Epidemiology and Detection Laboratory staff and students for conducting the antimicrobial susceptibility testing, serogrouping of the isolates, and PFGE analysis.
Disclosure Statement
No competing financial interests exist.