
Abstract
The major objective of this study was to investigate the prevalence of Shiga toxin (Stx)–producing Escherichia coli (STEC) in different types of food samples and to compare their genetic relatedness with STEC strains previously isolated from animal sources in Bangladesh. We investigated a total of 213 food samples, including 90 raw meat samples collected from retail butcher shops, 20 raw milk samples from domestic cattle, and 103 fresh juice samples from street vendors in Dhaka city. We found that more than 68% (n = 62) of the raw meat samples were positive for the stx gene(s); 34% (n = 21) of buffalo meats and 66% (n = 41) of beef. Approximately 10% (n = 2) of the raw milk and 8% (n = 8) of the fresh juice samples were positive for stx. We isolated STEC O157 from seven meat samples (7.8%), of which two were from buffalo meats and five from beef; and no other STEC serotypes could be isolated. We could not isolate STEC from any of the stx-positive raw milk and juice samples. The STEC O157 isolates from raw meats were positive for the stx2 , eae, katP, etpD, and enterohemorrhagic E. coli hly virulence genes, and they belonged to three different phage types: 8 (14.3%), 31 (42.8%), and 32 (42.8%). Pulsed-field gel electrophoresis (PFGE) typing revealed six distinct patterns among seven isolates of STEC O157, suggesting a heterogeneous clonal diversity. Of the six PFGE patterns, one was identical and the other two were ≥90% related to PFGE patterns of STEC O157 strains previously isolated from animal feces, indicating that raw meats are readily contaminated with fecal materials. This study represents the first survey of STEC in the food chain in Bangladesh.
Introduction
S
The pathogenicity of STEC strains is associated with various virulence factors. The main factor is the capacity to forms two potent phage-encoded cytotoxins called Shiga toxins (Stx1, Stx2, and their variants) or VT1 and VT2 (Paton and Paton, 1998). Another virulence factor associated with clinical STEC isolates is the protein intimin, encoded by the eae gene and responsible for the intimate attachment of STEC strains to the intestinal epithelial cells and causing the formation of attaching and effacing lesions in the intestinal mucosa (Kaper et al., 1998). Enterohemolysin, also called enterohemorrhagic E. coli (EHEC) hemolysin has been identified as a possible STEC virulence factor (Beutin et al., 1989). Two other virulence genes, katP and etpD, located in the large plasmid are also found in human pathogenic strains of E. coli O157. katP encodes the bifunctional catalase peroxidase, which protects the strains from oxidative stress exerted by host immune cells (Brunder et al., 1996). The etpD gene cluster is closely related to type II secretion pathway operons of Gram negative bacteria (Schmidt et al., 1997).
STEC strains rarely cause disease in animals, and ruminants are recognized as their main natural reservoir. Cattle are considered to be the major animal source of STEC that are virulent to humans, in particular STEC O157 (Caprioli et al., 2005). Contamination of carcasses with STEC usually occurs during slaughter and subsequent processing through fecal material originating directly or indirectly from the rectal-cecal area (Erickson and Doyle, 2007). Dairy products (milk, cheese, and cream) associated with infection have included those that are unpasteurized, have had a pasteurization failure, or have been contaminated postpasteurization. Ready-to-eat foods have also been associated with infection, particularly cooked meats contaminated by raw materials during processing, in catering establishments, at retail sale, and in the home. In developed countries, STEC O157 outbreaks associated with beef have caused concern among public health workers, clinicians, and the public, prompting major changes in clinical and laboratory practice, meat production, and food preparation (Mahon et al., 1997).
Similar to many of the developing and underdeveloped countries, the hygienic conditions in Bangladesh are severely compromised, and living with domestic animals within the same premises is a common practice among the Bangladeshi population. In our previous study, we found that the prevalence of STEC in slaughter animals was 24% and 43% of these STEC isolates belonged to serogroup O157 (Islam et al., 2008). This high prevalence of STEC in animal reservoirs appears to have a little impact on public health, considering the lack of STEC-associated infections among the Bangladeshi population (Islam et al., 2007). Nevertheless, to determine the underlying risk of infection with STEC among the Bangladeshi population, it is important to know the occurrence and transmission of these organisms in the human food chain.
Since the incidence of STEC in Bangladeshi foods has remained unknown, the study being reported here examined the occurrence of STEC in different types of food samples, with the isolates further characterized according to serotype, phage type (PT), virulence gene profile, and pulsed-field gel electrophoresis (PFGE) type. In addition, we compared the PFGE types of the food isolates with that of a collection of STEC strains isolated at adjacent period from animal feces in Bangladesh to determine clonal relatedness.
Materials and Methods
Sampling
We randomly collected 90 raw meat samples (60 beef and 30 buffalo) from July 2006 to January 2007 from 15 retail butcher shops located in five different areas (3 from each area) in Dhaka city. From each butcher shop, we collected six meat samples at two different time points (three at one time point) during the sampling period. In the same period, we randomly collected raw milk samples from 20 dairy cows domesticated at 20 different households located in two suburbs (10 from each area) of Dhaka city. In addition, we collected 103 freshly prepared juice samples, including mixed herbal juice (n = 49), sugar cane juice (n = 40), wood apple juice (n = 7) and some other types of juice (three milk shake, one lemon, two mix fruit, and one pineapple) from street vendors who operate in the main streets of 10 different locations in Dhaka city. All types of juice were freshly extracted at the vendor premises and did not constitute dilution from a proprietary concentrate. Only one juice sample was collected at one time point from each vendor. Each sample (minimum of 25 g or 25 mL) was aseptically transferred to a sterile plastic bag, placed in an ice box, and transported to the laboratory within 2 h. We started microbiological investigation of the samples within 4 h after collection.
Isolation of STEC O157
For meat samples, a 25-g portion, and for raw milk and juice samples, a 25-mL portion was added to 225 mL of modified tryptone soy broth (Oxoid, Basingstoke, United Kingdom). After homogenization in a stomacher for 1 min, the samples were incubated for 18–20 h at 37°C. STEC O157 strains were isolated as previously described (Islam et al., 2006), by using the immunomagnetic separation technique; and presumptive isolates were confirmed by an agglutination test using E. coli O157 latex agglutination kit (Murex Biotech, Dartford, Kent, United Kingdom) and a polymerase chain reaction (PCR) for the rfb (O-antigen-encoding) region of E. coli O157 (rfbEO157) (Paton and Paton, 1998).
Isolation procedure for STEC non-O157
The overnight enrichment cultures were screened by PCR for the presence of stx 1 and stx 2 using the primers as described earlier (Islam et al., 2007). DNA was extracted from 1 mL of broth culture by thermal cell lysis using Chelex-100 resin (Bio-Rad Laboratories, Richmond, CA), and 5 μL of the DNA extract was used in the PCR (Malorny et al., 2003). Multiplex PCR for stx 1 and stx 2 was carried out in a PTC-200 peltier thermal cycler (Bio-Rad) using the program as previously described (Islam et al., 2007). DNA extracted from E. coli O157:H7 strain NCTC 12079 and E. coli strain ATCC 25922 were used as positive and negative controls, respectively, in each PCR reaction. Broth cultures that yielded positive PCR results for stx 1 and/or stx 2 were streaked onto tryptone bile x-glucuronide (TBX) agar (Oxoid) supplemented with cefixime (20 mg/L), cefsulodin (3 mg/L), and vancomycin (30 mg/L) (CCV-TBX). Single colonies, including different morphological types, were transferred from the CCV-TBX and plated onto Luria agar (Becton Dickinson, Sparks, MD) to create a grid pattern of 96 colonies (12 × 8). The plates were incubated overnight at 37°C. The isolates were subjected to PCR for the detection of stx 1 and stx 2 as described above. The number of PCR reactions was reduced to 20 by pooling the colonies per row and per column in 500 μL sterile distilled water. DNA was extracted by boiling the suspensions for 10 min, and 5 μL of the supernatant was used in the PCR. The isolates with positive PCR results were identified as E. coli by an API 20E test (bioMérieux, Marcy l'Etoile, France).
PCR detection of virulence factors
PCR for detecting both chromosome (e.g., stx 1, stx 2, and eae)- and plasmid (e.g., hly EHEC, katP, and etpD)-encoded virulence genes was performed as described earlier (Islam et al., 2007) in a total volume of 25 μL containing 2.5 μL of 10 × Fast Start buffer with 1.5 mM MgCl2 (Applied Biosystems, Weiterstadt, Germany), 200 μM concentration of each deoxynucleoside triphosphate, a 0.2 pmol of each primer, and 1 U of Fast start Taq DNA polymerase (Roche, Mannheim, Germany) using a PTC-200 peltier thermal cycler (Bio-Rad).
Phage typing
Isolates confirmed to be STEC O157 were phage typed at the laboratory of Enteric Pathogens, Health Protection Agency Centre for Infections, London, United Kingdom.
Pulsed-field gel electrophoresis
PFGE was performed following the standardized protocol developed by PulseNet for E. coli O157:H7 (Ribot et al., 2006). Analysis of the TIFF images was carried out by the BioNumerics software (Applied Maths, Sint-Martens-Latem, Belgium) using the dice coefficient and unweighted-pair group method using average linkages to generate dendrograms with 1.0% tolerance values. Normalized PFGE patterns (bundle files) of the food isolates were compared with those of the animal isolates from our previous studies (Islam et al., 2008) to determine the clonal relationships.
Statistical analysis
Data were entered and statistical analyses were performed using Statistical Package for the Social Sciences version 11.5. The significance of difference was evaluated by the χ2 test, and Fisher's exact test was applied when appropriate. A p-value of <0.05 was considered statistically significant.
Results and discussion
Occurrence of STEC in food samples
Of the 90 meat samples tested in this study, we found that 62 (69%) were positive for stx gene(s); 21 samples of buffalo meats and 41 samples of beef (Table 1). However, this difference was not statistically significant (p > 0.10). STEC O157 strains were isolated from seven samples (7.8%); two samples of buffalo meats and five samples of beef (Table 1). No STEC non-O157 strains were isolated. During the processing of carcasses, meat can easily become contaminated with animal feces if there are unhygienic conditions, lack of good processing practices, and a lack of knowledge of the butchers on basic hygiene. These features are commonly observed in butcher shops in Bangladesh, where the same premise is being used to slaughter animals, to process carcasses, and as vending point. Our result correlates with data reported from neighboring countries, including India, where STEC O157 have been identified as one of the predominant serogroups from buffalo meats (Hazarika et al., 2007). STEC O157 strains are isolated from raw minced beef samples (9%; n = 22) (Dutta et al., 2000), beef surface swabs (3.7%; n = 27), and milk samples (2.4%; n = 81) (Manna et al., 2006) in India. Of the 20 raw milk samples, two (10%) were positive for stx gene(s) (Table 1). Of the 103 fresh juice samples, eight (8%) were positive for stx gene(s); six samples of mixed herbal juices and two samples of wood apple juices (Table 1). Statistical analysis of data revealed that a significantly high number of raw meat samples were positive for stx gene(s) when compared with raw milk and street vended juice samples (p < 0.001). No STEC could be isolated from stx-positive samples of raw milk and street vended juices. This might be due to the presence of a very small number of organisms present in the sample that were insensitive to the culture method used in the study. Another reason might be the presence of free stx phages in the sample that turned out to be positive by PCR for stx genes.
By using immunomagnetic separation.
PCR, polymerase chain reaction.
Characterization of STEC O157 isolates
STEC O157 strains that were isolated from raw meat samples were further characterized. All isolates were positive for the stx 2 gene only. There is considerable epidemiological evidence to indicate that STEC O157 isolates producing Stx2 are more commonly associated with serious disease than isolates producing Stx1or Stx1and Stx2 (Law, 2000). In addition to the stx 2 gene, all isolates in the study were positive for eae, etpD, katP, and hly EHEC genes, which accentuate their capability to cause human infection.
The strains could be grouped into three different PTs: PT 31 (n = 4), PT 32 (n = 2), and PT 8 (n = 1). PTs 31 and 32 have been commonly found among STEC O157 isolates obtained from both human and nonhuman sources (food and animal feces) from different countries, suggesting a foodborne transmission of the organism (Khakria et al., 1990; Mannix et al., 2007). In our previous study, we found that PT 14 and PT 31 were the predominant PTs followed by PT 54 and PT 32 among STEC O157 strains isolated from animal feces (Islam et al., 2008). In this study, we did not find that any of the isolates belonged to PT14. PT 8 has been frequently found among STEC O157 strains from humans and bovine in many European countries, including Spain, Belgium, Finland, Germany, Italy, England, and Scotland (Khakria et al., 1990). We also found this PT in our previous study in an STEC O157 strain isolated from goat feces. Therefore, it is more likely that raw meat products are often contaminated with animal feces in the butcher shop environment in Bangladesh.
PFGE analysis of the seven STEC O157 isolates revealed six distinct patterns, suggesting a genetically heterogeneous group. We compared these patterns with a panel of PFGE pattern that comprises 44 STEC O157 strains previously isolated from feces of cows, buffalos, and goats from slaughterhouses in Dhaka city (Fig. 1). One strain (CM52) isolated from beef had an identical PFGE pattern with that of a strain isolated from buffalo feces (M139). Interestingly, both the beef and fecal strains were isolated from the same slaughterhouse (data not shown). Another beef isolate (CM2) had a PFGE pattern that was 92% related to that of a cluster of strains isolated from cow (C8) and goat feces (G72 and G85). Two strains, one from beef (CM56) and one from buffalo meat (MM28), belonged to the same clonal type (Fig. 1); and this PFGE pattern was 90% related to that of a strain previously isolated from cow feces (C21). Among the six PFGE patterns obtained from meat isolates, three (50%) were identical or closely (≥90%) related to those of the isolates from animal feces. However, the PFGE typing results did not correlate with the phage typing results. Overall results of the study indicate that meats are readily contaminated with fecal materials containing STEC O157 in the butcher shops due to unhygienic practices of carcass processing.
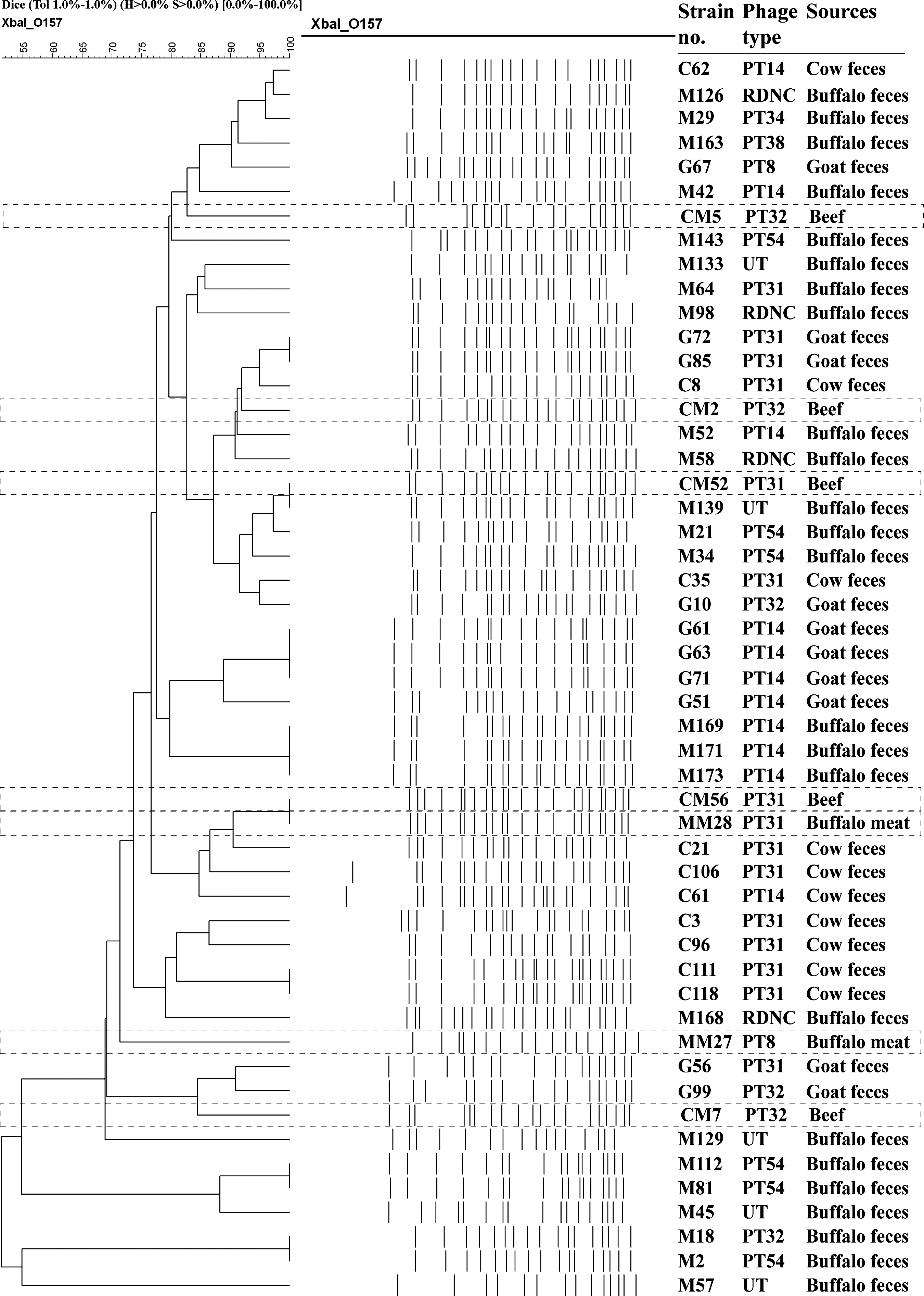
Dendrogram generated by Bionumeric software, showing distance calculated by the dice similarity index of pulsed-field gel electrophoresis XbaI profiles for STEC O157 isolates isolated from raw meats (n = 7) and animal feces (n = 44). Box with dotted line indicated profiles of raw meat isolates. The degree of similarity (%) is shown on the scale.
Conclusion
To our knowledge, this is the first survey to establish the prevalence of STEC in various food items in Bangladesh. Despite frequent contamination of raw meat products by potentially virulent E. coli O157, to date, no clinical data about impact on human health are available in Bangladesh. Improved surveillance and hygienic conditions along with the development of control measures are, therefore, highly recommended to minimize contamination of raw meat products by potentially virulent E. coli O157.
Footnotes
Acknowledgments
This research was funded by the International Centre for Diarrhoeal Disease Research, Bangladesh (ICDDR,B) and by the Netherlands Foundation for the Advancement of Tropical Research (NWO-WOTRO) grant (Award number WB 93-415). ICDDR,B acknowledges with gratitude the commitment of NWO-WOTRO grant to the Center's research efforts. ICDDR,B also gratefully acknowledges these donors who provide unrestricted support to the Center's research efforts: Australian International Development Agency, Government of Bangladesh, Canadian International Development Agency, The Kingdom of Saudi Arabia, Government of the Netherlands, Government of Sri Lanka, Swedish International Development Cooperative Agency, Swiss Development Cooperation, and Department for International Development, United Kingdom.
Disclosure Statement
No competing financial interests exist.