
Abstract
In the present study, the bacteriocins produced by Staphylococcus aureus 4185, a strain isolated from bovine mastitis, were purified and partially characterized. After purification by ammonium sulfate precipitation, cation-exchange chromatography, and five runs of high-performance liquid chromatography (HPLC), antimicrobial activity was recovered with 40% and 80% isopropanol, suggesting that more than one antimicrobial peptide, named aureocins 4185, is produced by S. aureus 4185. Mass spectrometry analyses revealed three peptides eluted with 40% isopropanol: peptide A (2,305.3 ± 1.5 Da), peptide B (2,327.3 ± 1.5 Da), and peptide C (3,005.5 ± 1.5 Da), and two peptides eluted with 80% isopropanol: peptide D (6,413.5 ± 1.5 Da) and peptide E (12,834.5 ± 1.5 Da). Although five peptides have been detected, only four small peptide sequences were obtained by matrix-assisted laser desorption/ionization time of flight (MALDI-TOF)/TOF mass spectrometry analyses: SLLEQFTGK (eluted with 40% isopropanol), ALLYDER, NNTSHNLPLGWFNVK, and NNLAQGTFNATK (eluted with 80% isopropanol). The sequences SLLEQFTGK and ALLYDER revealed identity with hypothetical peptides with unknown function. The sequences NNTSHNLPLGWFNVK and NNLAQGTFNATK showed similarity to a segment of a precursor of staphylococcal autolysins. The antimicrobial activity detected in the supernatant of strain 4185 proved to be resistant to heat treatment at 65°C; however, treatment at 80°C abolished completely its antimicrobial properties. The concentrated supernatant containing aureocins 4185 exhibited a strong bacteriolytic activity toward Micrococcus luteus ATCC 4698. Additionally, aureocins 4185 exhibited antagonistic activity against important foodborne pathogens, including Listeria monocytogenes, thus showing a potential application in food preservation.
Introduction
L. monocytogenes is an important food pathogen, accounting for a high mortality rate (Swaminathan and Gerner-Smidt, 2007; Sergelidis and Abrahim, 2009). B. cereus is a spore-forming food-poisoning bacterium, frequently isolated from a remarkable range of foods (Stenfors-Arnesen et al., 2008). S. aureus is another important pathogen transmitted by food, being involved in numerous outbreaks in several countries (Murray, 2005).
The demand for high-quality, “natural,” “minimally processed,” and safe foods has been growing. To avoid the transmission of undesirable microorganisms, while satisfying the consumers demand, is the major challenge for the food industry. A promising alternative to control food pathogens and food spoilage bacteria could be the use of bacteriocins (Gálvez et al., 2007; Bastos and Ceotto, 2010).
Bacteriocins are antimicrobial peptides with inhibitory activity against other bacteria, which offer attractive options for food preservation. The use of bacteriocins as biopreservatives in food systems have some advantages such as follows: (i) it could decrease the risk for transmission of foodborne pathogens through the food chain and ameliorate the economic losses due to food spoilage, (ii) it may permit the application of less severe physical–chemical treatments without compromising food safety, and (iii) it could reduce the use of chemical preservatives (Gálvéz et al., 2007; Bastos and Ceotto, 2010).
The bacteriocins produced by Gram-positive bacteria generally have a broad spectrum of activity against foodborne pathogenic bacteria. Some bacteriocins, especially the lantibiotics, exhibit resistance to pH and heat. Additionally, they are inactive and nontoxic on eukaryotic cells (Gálvéz et al., 2007; Bastos and Ceotto, 2010).
At present, only nisin and pediocin PA-1/AcH are commercially used in food preservation. Nisin is commercially available as Nisaplin (Danisco) and pediocin PA1 has also been commercially used in the form of ALTA™ 2431-Quest (Cotter et al., 2005). Although enterocin AS-48 is not commercially available yet, it also presents a potential application in food systems (Bastos and Ceotto, 2010).
In a previous study (Ceotto et al., 2009), among several strains of S. aureus involved in bovine mastitis in Brazil, we have identified one strain, 4185, with antimicrobial activity against strains of Listeria spp. (including L. monocytogenes) and B. cereus, which are important foodborne pathogens. Therefore, in the present work, the antimicrobial substances produced by S. aureus 4185 were purified and characterized.
Materials and Methods
Bacterial strains and culture conditions
The bacteriocin-producing strain S. aureus 4185 was isolated from a bovine mastitis case in the southeast region of Brazil (Ceotto et al., 2009). Micrococcus luteus ATCC 4698, which is sensitive to staphylococcins, was used as an indicator in the inhibition assays (Ceotto et al., 2009). Both strains were cultivated in GM17 (M17 [Difco] supplemented with 5% [w/v] glucose) or tryptone soy broth (TSB) (Difco), at 37°C for 18 h. They were stored in TSB with 40% glycerol (w/v) at −20°C, until use. When necessary, the media were supplemented with agar, at either 0.6% (w/v) or 1.5% (w/v).
Bacteriocin activity assays
The inhibitory action of bacteriocin was investigated either on agar plates, by the deferred-antagonism technique (Giambiagi-DeMarval et al., 1990), or by the serial dilution method in microtiter plates, as previously described (Ceotto et al., 2009).
Briefly, deferred-antagonism test was performed by spotting the producer strain, S. aureus 4185, on 20 mL of brain heart infusion (BHI) agar plates (1.5%, w/v). After 18 h at 37°C, 3 mL of BHI soft agar (0.6%, w/v), containing around 107 cells of the indicator strain, was sprayed over the plates. The plates were incubated under the same conditions and inhibition zones around the producer spots were measured in millimeter. The experiments were repeated at least three times. The indicator strains were considered sensitive to bacteriocin when it exhibited a clear inhibition zone.
Bacteriocin activity was also determined in microtiter plate assays. Each well of the microtiter plate contained 100 μL of double serial dilutions of the bacteriocin in GM17 medium and 100 μL of the diluted culture of the indicator strain (corresponding to an OD600 of 0.05). The plates were incubated at 37°C for 24 h and the growth was measured spectrophotometrically at 600 nm with a microtiter plate reader (Elx800; Biotek Instruments). The antimicrobial activity was defined as the highest dilution required to produce 50% growth inhibition of the indicator strain, compared with the control culture with no bacteriocin added, and it was expressed in bacteriocin units (BU)/mL.
Thirty-one different Gram-positive bacteria from our laboratory collection were used as indicators for the confirmation of the spectrum of activity: B. cereus I56, B. cereus 2710, Bacillus coagulans UH, Bacillus licheniformis ATCC 12759, Geobacillus stearothermophilus ATCC 7953, L. monocytogenes 11LM, L. monocytogenes L1/2A, L. monocytogenes 7898, 4 Listeria spp., and 19 Staphylococcus spp. strains isolated from raw vegetables. The experiments were repeated at least three times.
Purification of aureocin 4185
The bacteriocin was purified from 1 L culture of S. aureus 4185, grown using the best conditions for bacteriocin production, previously determined: in GM17 medium, at 37°C for 18 h, under shaking (180 rpm) (unpublished data). The culture was centrifuged at 12,000 g for 20 min at 4°C. Ammonium sulfate (40%, w/v; Merck) was added to the supernatant, which was kept under shaking for 2 h at 4°C. The bacteriocin was precipitated from the supernatant by centrifugation at 12,000 g for 20 min at 4°C and dissolved in 100 mL sterile Milli-Q water. The solution pH was adjusted to 4.0 with 1 M HCl (Merck) and subsequently subjected to a cation-exchange column (SP Sepharose Fast Flow; Amersham Biosciences) previously equilibrated with 10 mM acetic acid (Merck). The cation-exchange column was washed with 100 mL of 10 mM sodium phosphate buffer (pH 6.8) before the elution of bacteriocin with 1 M NaCl. The purification was completed by at least two consecutive runs of HPLC, using an Äkta Purifier System (Amersham Pharmacia Biotech). Initially, the sample was applied onto the Resource RPC 1-mL column (Amersham Pharmacia Biotech), followed by purification in a Sephasil Peptide C8 5-μm ST 4.6/250 column (Amersham Pharmacia Biotech). When necessary, the fractions with the highest antimicrobial activity and eluted from the second HPLC run were subjected to a third run in a Sephasil Peptide C8 5-μm ST 4.6/250 column. Elution was performed with a water–isopropanol gradient from 0% to 100% (v/v; Merck), containing 0.1% trifluoroacetic acid (TFA) (v/v; Merck), and it was monitored at 214 nm. The fractions showing the highest bacteriocin activity were used in the subsequent purification step.
Mass spectrometry analyses and N-terminal amino acid sequencing
The molecular mass of the purified peptides was determined by matrix-assisted laser desorption/ionzation time of flight (MALDI-TOF) mass spectrometry, with a matrix-assisted laser desorption ionization-time of flight mass spectrometer (Voyager-RP DE; Applied Biosystems) in the linear positive-ion mode, with a total acceleration voltage of 25 kV, as described by Brede et al. (2004). The amino acid sequence was determined by MALDI-TOF/TOF spectrometry, as described by Lee et al. (2001) and Peters et al. (2004).
Determination of the bacteriocin activity kinetics
This experiment was performed by reading the optical density (in a Bioscreen C equipment; Oy Growth Curves AB Ltd.) of the indicator culture in the presence of the bacteriocin, for 8 h at 37°C. The partially purified bacteriocin (1024 BU), obtained by ammonium sulfate precipitation followed by cation-exchange chromatography, was tested against 109 cells of the indicator strain, M. luteus ATCC 4698, in a 400 μL final volume. The OD600 was measured in intervals of 15 min.
Effects of temperature on bacteriocin activity
S. aureus 4185 was cultivated in 100 mL GM17 medium, at 37°C for 18 h, under shaking (180 rpm). The culture was centrifuged at 12,000 g for 20 min at 4°C, and the supernatant was filtered (0.25 μm filters; Sarstedt) and treated at 65°C and 80°C for 15 min. After treatment, the residual activity of each sample was compared with that of the unheated control by microtiter plate assays.
Results and Discussion
Bacteriocin purification
The bacteriocins were purified from 1 L culture of S. aureus 4185 cultivated in the best conditions for bacteriocin production. After bacterial growth, the culture supernatant was subjected to ammonium sulfate precipitation and subsequently to cation-exchange chromatography and 256,600 BU were eluted in 30 mL of 1 M NaCl. The purification was completed by five runs of HPLC. Initially, the sample was applied onto the Resource RPC 1-mL column and 24 fractions of 1 mL were recovered. Antimicrobial activity was detected in fractions 13–19, with a total of >100,000 BU. Fractions 14 and 17 had >20,480 BU, each. Fractions 13 and 14 (>52,000 BU), obtained in the first HPLC, were designated as sample 1. Fractions 15–19 (>60,000 BU), obtained in the first HPLC, were designated as sample 2. Both samples 1 and 2 were separately subjected to a second run of HPLC, using the Sephasil Peptide C8 column, and the fractions showing the highest bacteriocin activity were used in a third run of HPLC.
After two runs of HPLC in a Sephasil Peptide C8 column, 23,360 BU (final fractions 9–11 [Fig. 1]), eluted with 40% isopropanol, were recovered from sample 1. From sample 2, 61,440 BU, present in fractions 13 and 14 (Fig. 2), were recovered with 80% isopropanol. These results suggest that the antimicrobial activity found in the supernatant of strain 4185 was related to the presence of at least two different peptides. Like most staphylococcins (Bastos et al., 2009), these peptides proved to be cationic and hydrophobic, especially the peptides eluted with 80% isopropanol. The final fractions were used to determine the molecular mass and the amino acid sequences.
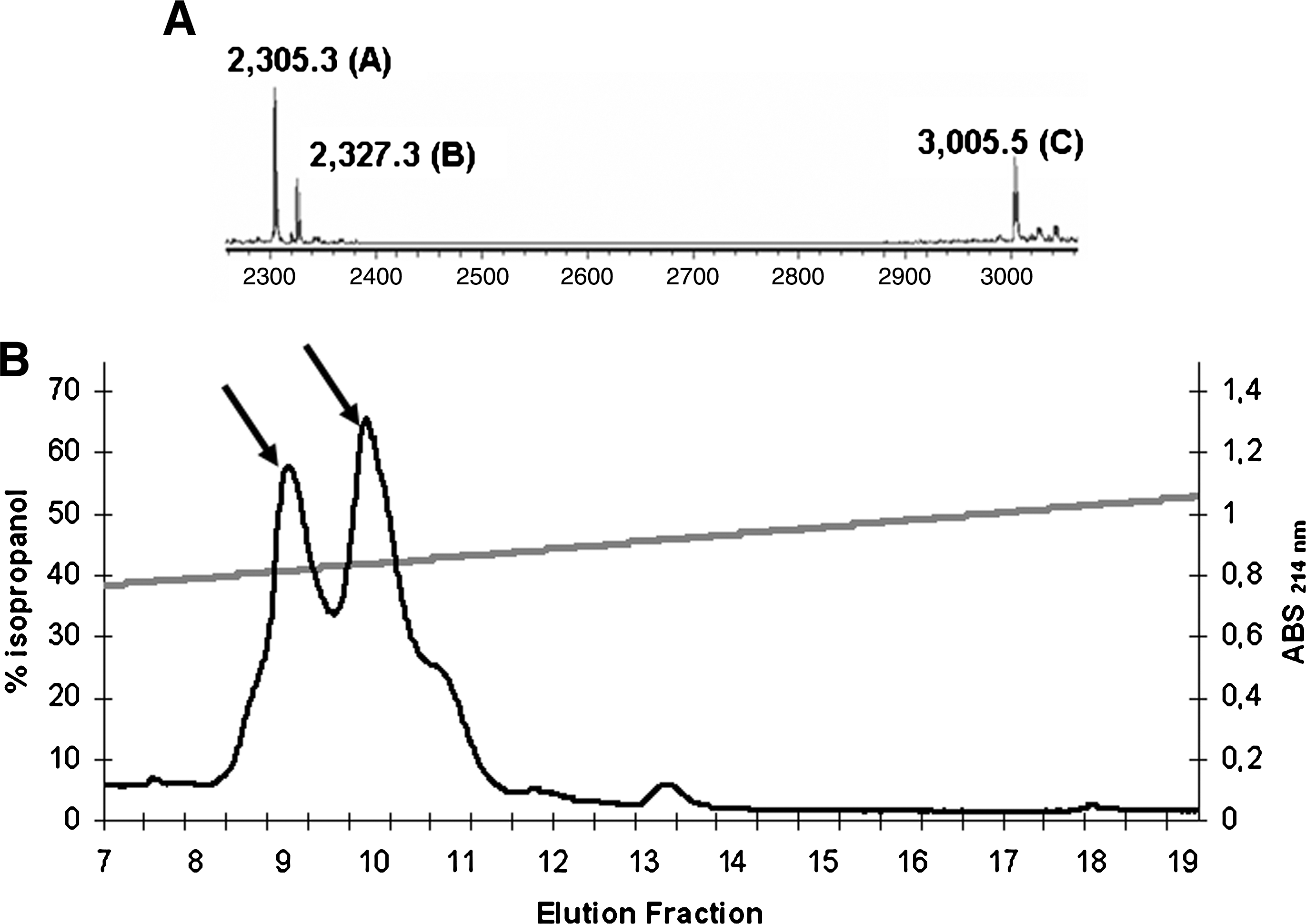
Aureocins 4185 purification by high-performance liquid chromatography (HPLC) and molecular mass determination (fractions eluted in 40% isopropanol). (
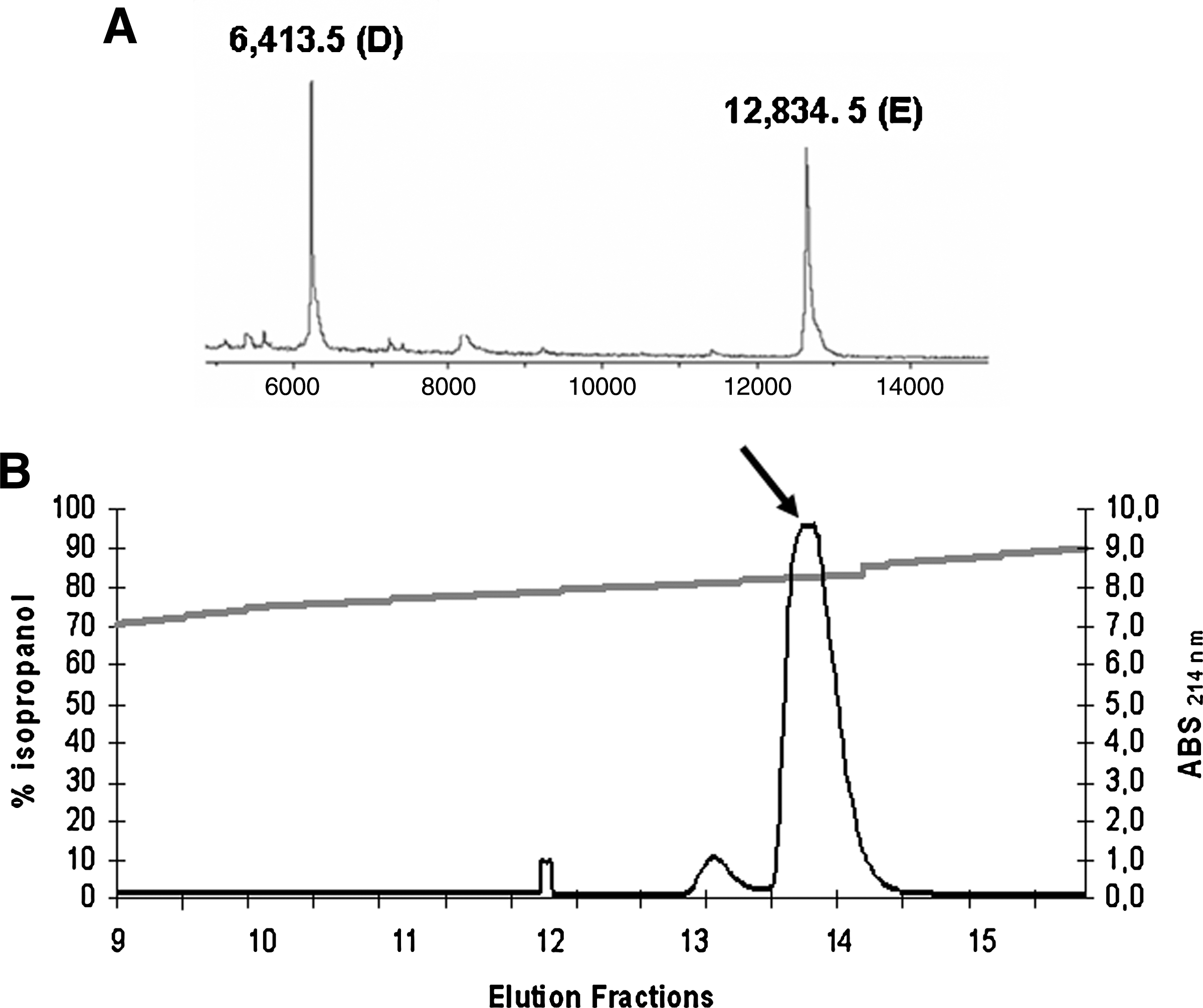
Aureocins 4185 purification by high-performance liquid chromatography (HPLC) and molecular mass determination (fractions eluted in 80% isopropanol). (
Molecular characterization of aureocins 4185
The determination of the molecular mass of the purified peptides revealed the presence of five different components. The M r of the peptides contained in the final fractions 9–11, eluted with 40% isopropanol (Fig. 1), was 2,305.3 ± 1.5 Da (peptide A), 2,327.3 ± 1.5 Da (peptide B), and 3,005.5 ± 1.5 Da (peptide C). The molecular mass of the peptides present in the final fractions 13 and 14, eluted with 80% isopropanol (Fig. 2), was 6,413.5 ± 1.5 Da (peptide D) and 12,834.5 ± 1.5 Da (peptide E).
Only four small peptide sequences were determined by MALDI-TOF/TOF mass spectrometry analyses: SLLEQFTGK from sequencing of the peptides contained in the fractions 9–11 (eluted with 40% isopropanol); ALLYDER, NNTSHNLPLGWFNVK, and NNLAQGTFNATK from sequencing of the peptides present in fractions 13 and 14 (eluted with 80% isopropanol). These results are summarized in Table 1.
In silico analysis of the sequence SLLEQFTGK revealed a high similarity with the C-terminal region of a 21-amino acid hypothetical peptide, MGIIAGIIKVIKSLIEQFTGK [theoretical M r of 2,259.82 Da, calculated by the program Compute pI/Mw (ExPASy Proteomics Server)], found in S. aureus USA300. This hydrophobic and cationic (theoretical pI of 9.7) hypothetical peptide showed no similarity to any protein deposited in the GenBank database.
From the fractions eluted with 80% isopropanol, three partial sequences of amino acids were obtained. The first sequence, ALLYDER, showed similarity with a fragment of a hypothetical protein of S. aureus RF122, with 144 amino acids. This hypothetical protein does not have a known function, but in silico analysis using the program “Blast Conserved Domains” (NCBI) revealed that it has a conserved domain with the mitogen-activated protein (MAP) proteins, enzymes involved in intracellular signaling (Gotoh and Nishida, 1998). The M r of this hypothetical protein was estimated to be ∼15.8 kDa. As a pretreatment with trypsin had been performed with the fractions exhibiting antimicrobial activity before amino acid sequencing, the fragments generated by digestion of this hypothetical protein with trypsin were also analyzed using the aforementioned program. None of the fragments generated would have a M r of either 6.4 or 12.8 kDa, which are the masses of peptides D and E. Therefore, it is unlikely that any of these peptides is related to this hypothetical protein.
Two other sequences, NNTSHNLPLGWFNVK and NNLAQGTFNATK, showed similarity to a segment of a precursor of an autolysin with 1245 amino acids, found in S. aureus Newman. The autolysins described in the literature are large proteins (50–100 kDa) that act on the cell wall of Gram-positive bacteria (Vollmer et al., 2008). The predicted M r and pI of this autolysin are 137.39 and 9.62 kDa, respectively (calculated by Compute pI/Mw). However, no large protein with antimicrobial activity was found in the fractions eluted with 80% isopropanol. Only two components, peptide D (6,413.5 ± 1.5 Da) and peptide E (12,834.5 ± 1.5 Da), were found in these fractions. Therefore, it is unlikely that any of the purified antimicrobial substances is an autolysin. Interestingly, the larger peptide E has a mass corresponding to twice the mass of peptide D. Peptide E could be a dimer of peptides D, or alternatively, peptide D could be a product of cleavage of peptide E.
Although most Bac+ strains produce only a single bacteriocin (Bastos et al., 2009), some bacterial strains produce more than one bacteriocin (Quadri et al., 1997). It is also possible that one of the bacteriocins produced by strain 4185 is composed of more than one peptide. Multicomponent bacteriocins have already been described in the literature, such as aureocin A70, which is composed of four peptides (Netz et al., 2001), and staphylococcin C55 (Navaratna et al., 1998) and lactococcin G (Nissen-Meyer et al., 1992), which are both composed of two peptides.
Effects of temperature on bacteriocin activity
The bioactive peptides produced by S. aureus 4185 seem to be resistant to treatment at 65°C for 15 min, because no change in activity (160 BU) was observed in both filtered (control) and heated supernatants. However, the treatment at 80°C for 15 min abolished completely the antimicrobial activity toward M. luteus, suggesting that the bacteriocins are sensitive to this temperature.
The relative heat sensitivity of these peptides suggests that they are not lantibiotics. Lantibiotics are small peptide (<5 kDa) bacteriocins characterized by the presence of a high proportion of unusual amino acids, such as lanthionine and 3-methyl-lanthionine, among others (Bierbaum and Sahl, 2009). Lanthionine and 3-methyl-lanthionine possess thioether intramolecular bridges and, as a consequence of their presence, lantibiotics are polycyclic structures that exhibit a remarkable resistance to heat. Class III was also excluded because it is composed of large (≥25 kDa) proteins (Bastos et al., 2009). Therefore, the antimicrobial peptides produced by S. aureus 4185 should belong to class II. Class II bacteriocins are small (<10 kDa), unmodified peptides (Nissen-Meyer et al., 2009).
The only staphylococcal class II bacteriocins characterized so far are aureocin A70 (Netz et al., 2001) and aureocin A53 (Netz et al., 2002a). Aureocin A70 is a four-component bacteriocin and the molecular mass of its components varies from ∼2.8 to ∼3.1 kDa (Table 2). Aureocin A53 is composed of a single peptide whose molecular mass is 6.01 kDa.
All the data related to the initial characterization of the peptides produced by S. aureus 4185 have already been published (Ceotto et al., 2009). These studies included PCR amplification of bacteriocin structural genes and DNA hybridization analyses, which showed that these peptides are distinct from all staphylococcins characterized until 2009. Such findings were corroborated by the molecular masses of the peptides found in both fractions with antimicrobial activity, which were different from those of all known class II staphylococcins described to date (Table 2). Recently, a new staphylococcin named Bsa has been described (Daly et al., 2010). However, Bsa is a 2.09-kDa lantibiotic, related to epidermin. Taken together, these data suggest that the antimicrobial peptides produced by S. aureus 4185 may represent new staphylococcins and from hereafter they will be referred as aureocins 4185.
Activity kinetics
The activity kinetics of aureocins 4185 was determined in a Bioscreen C, against M. luteus ATCC 4698, using a partially purified bacteriocin preparation obtained by precipitation with ammonium sulfate followed by cation-exchange chromatography. The OD600 of the indicator culture was reduced from 1.5 to ∼0.5 in 4 h (Fig. 3), suggesting that these antimicrobial peptides have a lytic activity against M. luteus ATCC 4698. These results suggest that either one of the peptides or the peptides combined have a strong bacteriolytic activity.
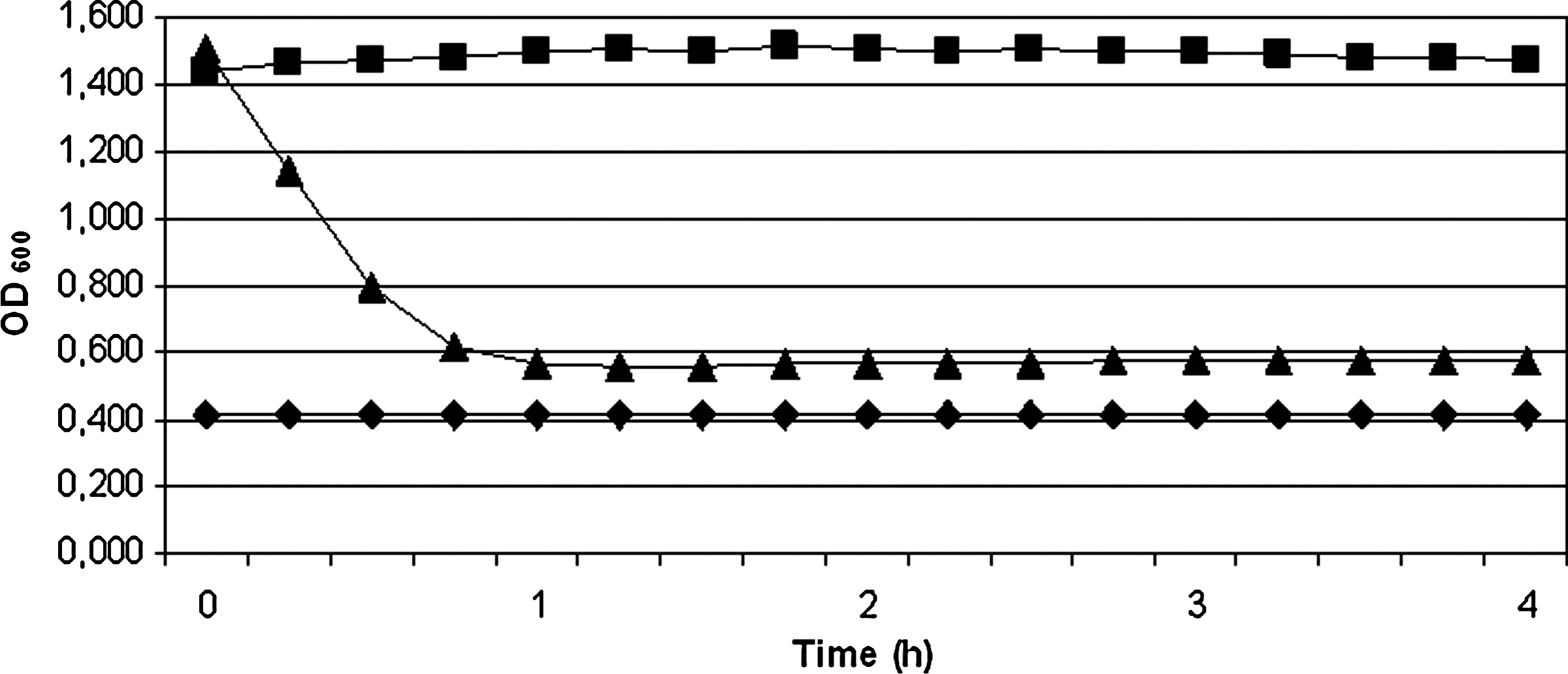
Aureocins 4185 activity kinetics. The experiment was performed in a Bioscreen C, by monitoring the optical density (OD600) of a Micrococcus luteus ATCC 4698 suspension, at every 15 min, for 4 h. (♦) GM17 medium (blank); (▪) M. luteus ATCC 4698 (control); (▴) M. luteus ATCC 4698 + aureocins 4185 (1024 BU). The results represent the average of two independent experiments.
Many bacteriocins have been shown to possess bacteriolytic activity. Lantibiotics, such as nisin (Cotter et al., 2005), and most class II bacteriocins, such as aureocin A53 (Netz et al., 2002b), act by permeabilization of the bacterial membranes and some class III bacteriocins, such as lysostaphin, act through the hydrolysis of the peptidoglycan of Gram-positive bacteria (Bastos et al., 2009). Although the individual mode of action of each aureocin 4185 has not been determined yet, it is possible that at least one of them is bacteriolytic.
Spectrum of activity
S. aureus 4185 showed antagonistic activity against B. cereus 2710, G. stearothermophilus, and one strain of Listeria spp. on agar plates. However, tests performed in microtiter plates showed that not only the filtered supernatant of S. aureus 4185 but also the supernatant treated at 65°C inhibited all the Bacillus (B. cereus I56, B. cereus 2710, B. coagulans, and B. licheniformis) and Listeria (L. monocytogenes 11LM, L. monocytogenes L1/2A, L. monocytogenes 7898, and four Listeria spp. isolated from raw vegetables) strains tested. Additionally, G. stearothermophilus and three (15.8%) out of 19 strains of Staphylococcus spp. were also inhibited in microtiter plate assays, suggesting that this method is more sensitive than the test performed on agar plates to detect antimicrobial activity.
S. aureus A70 Bac− supernatant was used as a negative control in these experiments. S. aureus A70 Bac− is the strain S. aureus A70 cured of pRJ6, which encodes aureocin A70 (Giambiagi-deMarval et al., 1990). The supernatant derived from the cured strain had no inhibitory activity. Therefore, neither the cultivation medium nor the staphylococcal nonbacteriocin products may account for the detected inhibitory activity.
Despite not showing a remarkable activity against staphylococci, aureocins 4185 had an inhibitory activity against Listeria spp. strains, including L. monocytogenes and listerial strains isolated from food, and B. cereus, two important foodborne pathogens. Additionally, B. coagulans, B. licheniformis, and G. stearothermophilus, which are food contaminants, were also inhibited.
Among the foodborne diseases described in the literature, listeriosis is of particular concern to the food industry and to susceptible consumers that include pregnant women, infants, immunocompromised individuals, and the elderly. Its agent, L. monocytogenes, can be transmitted through contact with hands, utensils, or contaminated food, especially foods that are eaten raw in salads (Carrasco et al., 2007; Cordano and Jacquet, 2009).
B. cereus is a spore-forming food-poisoning bacterium, widespread in nature and frequently isolated from a remarkable range of different foods and food ingredients, including rice, dairy products, spices, dried foods, and vegetables. It is also well adapted for growth in the intestinal tract of insects and mammals, where it may cause an emetic or a diarrheal type of food-associated illness. Because of its widespread presence and its ability to form heat-stable endospores in a relatively short time, B. cereus has been difficult to control (Stenfors-Arnesen et al., 2008).
Other Bacillus species are also very important to the food industry because the spores of these microorganisms are resistant to heat treatment and, after germination, cause food spoilage. B. licheniformis have been isolated from milk (Banykó and Vyletelová, 2009), salad (Meldrum et al., 2009), and seafood (Cabo et al., 2009). B. coagulans is a microorganism of relevance to industry of fruit juices, because of its ability to withstand acid and promote the deterioration of the product (Bevilacqua et al., 2009).
G. stearothermophilus is a thermophile widely distributed in soil, hot springs, and ocean sediment and is a cause of spoilage in food products because of its ability to withstand heat treatment to which the commercially sterile products are subjected (Head et al., 2008).
Conclusions
The present data suggest that the antimicrobial activity exhibited by S. aureus 4185 is due to production of more than one antimicrobial peptide, which seem to represent new staphylococcins. Aureocins 4185 could be either a multicomponent bacteriocin or these peptides may represent different aureocins. Both possibilities are currently under investigation.
Aureocins 4185 have potential biotechnological applications as biopreservatives to prevent either food spoilage or foodborne diseases. Broad-spectrum bacteriocins, such as aureocins 4185, could be used to inhibit high-risk bacteria in foods, such as L. monocytogenes, B. cereus, B. licheniformis, B. coagulans, and G. stearothermophilus.
Although aureocins 4185 have been shown to be sensitive to heat treatment at 80°C, they could be used in preservation of foods that are either consumed raw or minimally processed. The bacteriocins could be added to the foods in the form of a concentrate or incorporated into a matrix of a bioactive food packaging. In the case of milk, for example, the bacteriocin could be added after pasteurization (Gálvéz et al., 2007).
Many authors have already demonstrated that the bacteriocins produced by lactic acid bacteria are inactive and nontoxic on eukaryotic cells and become inactivated by digestive proteases, having little influence on the gut microbiota (Gálvéz et al., 2007). These bacteriocins have the status generally recognized as safe (GRAS). Aureocins 4185 were previously shown to be sensitive to proteolytic enzymes. However, their cytotoxicity should be tested in the future with each individual bacteriocin.
It would be interesting to test also if aureocins 4185 are sporicidal, but we need first to determine which peptide has activity against B. cereus and the other spore-forming bacteria inhibited, because the antimicrobial activity observed could be related to the action of more than one peptide. These experiments wait for the individual cloning of each aureocin 4185 gene cluster.
Studies about the properties of each individual aureocin 4185 and an investigation of all aspects related to bacteriocin biology, including the elucidation of their structure–function relationships, production, immunity, regulation, spectrum, and mode of action, are therefore required before considering the applications of aureocins 4185.
Footnotes
Acknowledgments
The authors thank Dr. M. Skaugen from the Norwegian University of Life Science for performing the MS and the peptide sequencing analyses. H.C. and P.C.F. were recipients of scholarships from CNPq/Brazil. This study was supported by grants from CNPq, FAPERJ, and PRONEX (to M.C.F.B.).
Disclosure Statement
No competing financial interests exist.