
Abstract
The aim of this study was to investigate the response of Campylobacter jejuni at single-cell level when exposed to different concentrations of chlorine dioxide (ClO2). The parameter of choice, intracellular pH (pHi), was determined by using fluorescence ratio imaging microscopy with a pH-sensitive, ratiometric 5(6)-carboxyfluorescein diacetate succinimidyl ester probe. In addition, the culturability expressed in colony counts was determined. Our results revealed that several subpopulations with different physiological states, as judged by their pHi, were created by ClO2 treatment. The greater the concentration of ClO2, the smaller the subpopulation of healthy cells with pHi > 6.8 and the smaller the colony count as determined on nonselective agar plates. ClO2 at concentrations (60 ppm) induced injuries that resulted in complete loss of culturability and adversely affected the ability to resuscitate under subsequent more favorable conditions. The presence of injured cells in food could present a risk for public health. Additional hurdles have to be included in food preservation to suppress the survival and recovery of injured cells.
Introduction
Chlorine dioxide (ClO2) is a powerful oxidizing and sanitizing agent, with a broad biocidal activity against bacteria, yeast, viruses, fungi, and protozoa (Gomez-Lopez et al., 2009). ClO2 has been lately considered as an alternative to chlorine, most frequently used as a biocide, since its reactions with organic or inorganic species do not usually lead to the formation of harmful toxic by-products (Ayyildiz et al., 2009). ClO2 can be used for the disinfection of drinking water, but also in food industry for decontamination of fresh produce, fish, and meat (Gomez-Lopez et al., 2009). In addition to the numerous methods of application, many factors affect the efficacy of ClO2, such as the concentration used, the exposure duration, and the properties and state of the food products (Gomez-Lopez et al., 2009). Determination of pHi using fluorescence ratio imaging microscopy (FRIM) is a valuable tool to assess the physiological state of microorganisms at the single-cell level upon exposure to stress (Smigic et al., 2009, 2010). The aim of this study was to examine the effect of different concentrations of aqueous ClO2 on the change of pHi in Campylobacter jejuni and test the difference in the responses between two different C. jejuni isolates. Additionally, our interest was to examine the effect of ClO2-induced injury (60 ppm) on the subsequent culturability of C. jejuni cells under favorable growth conditions.
Materials and Methods
Bacterial strains and growth conditions
C. jejuni LFMFP (Laboratory of Food Microbiology and Food Preservation, University of Gent, Belgium) 595 (bovine isolate, LMG 8841) and C. jejuni LFMFP 602 (human isolate) (Cools et al., 2003) were used in this study. The standard maintenance of stock culture and its propagation using Bolton broth into a working culture were done as described elsewhere (Smigic et al., 2009). Colonies grown on Colombia base agar (CBA) plates (Oxoid) supplemented with horse blood (E&O Laboratories) and incubated for 48 h at 37°C in a microaerophilic atmosphere were used for both enumerations and pHi determinations. Colonies used for pHi determination were stained with pH-sensitive fluorescence probe 5(6)-carboxyfluorescein diacetate succinimidyl ester (CFDA-SE; Molecular Probe, Inc.) (Smigic et al., 2009).
Fluorescence labeling and FRIM
C. jejuni colonies grown on CBA plates were harvested and stained according to the protocol previously described by Smigic et al. (2009). Harvested cells were resuspended in 980 μL citrate phosphate buffer (pH 7.0), in which 10 μL of 1 M glucose (Sigma-Aldrich) and 10 μL of 4.48 mM CFDA-SE (Molecular Probe, Inc.) were added. After 30 min of incubation at 37°C under microaerophilic conditions, stained cells were harvested by centrifugation (12,000 g, 2 min), resuspended in physiological salt solution (0.85% w/v NaCl), and kept on ice in the dark before exposure to aqueous ClO2 (see the ClO2 treatment section). For the immobilization of C. jejuni cells, microscopic glass slides were coated with bind silane (PlusOne; Amercham Biosciences) using a previously described protocol (Smigic et al., 2009).
The pHi of single cells was determined using the FRIM method (Budde and Jakobsen, 2000; Smigic et al., 2009). Fluorescence microscopy, determination of ratio value, and calibration curves were obtained essentially as described by Smigic et al. (2009). Briefly, CFDA-SE-stained cells were excited at 488 and 435 nm wavelengths and emission was recorded, through 515- to 565-nm band pass emission filter, on a cooled charge-coupled device camera. Ratio values R 488/435 were calculated by dividing the intensity of individual pixels on the 488 nm image by the intensity of corresponding pixels at 435 nm image. The background (regions without cells) was subtracted. In each experiment a minimum of 40 single cells was analyzed. For pH calibration, CFDA-SE-labeled cells were permeabilized by incubation in ethanol (63% v/v, 37°C, 30 min) and resuspended in citrate phosphate buffers with pH ranging from 5.5 up till 8.0, allowing pHi to equilibrate to external pH (pHex). A polynomial model fitted to the data was used to calculate pHi.
ClO2 treatment
ClO2 was prepared and stored under the conditions previously described by Smigic et al. (2010). Before each experiment, the concentration of the stock ClO2 solution was determined using an iodometric method. Appropriate ClO2 concentrations were diluted from the stock solution, using MilliQ water. Bacterial cell suspensions were exposed to different concentrations of ClO2. For that purpose, 0.1 mL of ClO2 solution (200, 400, 600, and 800 ppm) was transferred into a tube containing 0.9 mL of C. jejuni cells (∼ 108 CFU/mL) to obtain 20, 40, 60, and 80 ppm ClO2 concentrations, respectively. The respective pH values were 4.0, 3.7, 3.4, and 3.2. The test tubes used for ClO2 treatment were covered with aluminum foil to avoid the degrading effect of light on ClO2. After 2 min of the exposure, Bolton broth without supplement (organic matter) was added to the cell suspension to eliminate the residual activity of ClO2. Subsequently, cells were harvested by centrifugation (12,000 g, 2 min), resuspended in 1 mL Bolton broth (pH 6.0), and kept on ice in the dark before the plating and pHi determination. pH of the solution with resuspended cells was not additionally adjusted. In each case at least 40 individual cells were analyzed for pHi determination. To determine the colony count C. jejuni cultures were plated out on CBA (supplemented with horse blood) and incubated (48 h at 37°C, microaerophilic atmosphere). Control experiments were performed using MilliQ water in place of aqueous ClO2. Decontamination with ClO2 and control treatments were performed in two independent experiments, for both colony count and pHi determination.
Behavior of C. jejuni following the exposure to ClO2
To investigate the ability of ClO2-treated C. jejuni cells to recover from the stress, cell suspensions treated with 60 ppm ClO2 were incubated under favorable growth conditions. For that purpose, 0.1 mL of the treated suspension (treatment procedure was described in ClO2 treatment section) was transferred into the tube containing 10 mL fresh Bolton broth with pH 7.0. Tubes were incubated at 37°C under microaerophilic atmosphere, and samples were plated out on CBA plates (supplemented with horse blood) immediately after the inoculation, and after 24 and 48 h of incubation.
Results and Discussion
Values of measured pHi and colony counts (log CFU/mL) obtained for two C. jejuni strains, 595 and 602, when treated with different ClO2 solutions (20, 40, 60, and 80 ppm), MilliQ water as a control, and untreated cultures (Bolton broth pH 6.5), are presented in Figure 1. The observed effect can be attributed to the ClO2 molecule as the pure effect of low pH has been previous found to be negligible (Smigic et al., 2009). For both C. jejuni strains, all cells in untreated culture (suspended in Bolton broth pH 6.5) had pHi higher than 6.8. When the cells were exposed to MilliQ water, the major subpopulation of the cells being 78% and 100% for C. jejuni 595 and 602, respectively, consisted of healthy and injured cells able to maintain a transmembrane proton gradient (pHi higher than pHex). At ClO2 concentrations of 20 and 40 ppm, this subpopulation of the cells decreased in both C. jejuni strains, but to a different extent. The subpopulation of the cells with positive pH gradient ranged from 55% to 70% for C. jejuni 595, whereas this was in the range from 90% to 95% for C. jejuni 602. The level of reduction for C. jejuni 595 when treated with 20 or 40 ppm ClO2 was 1.4 and 2.9 log CFU/mL, respectively, when compared to water treatment (0 ppm ClO2). However, in more ClO2-resistant strain, C. jejuni 602, the same treatments of 20 and 40 ppm ClO2 only caused reductions of 1.1 and 1.9 log CFU/mL, respectively. Exposure of C. jejuni cells to increasing ClO2 concentrations resulted in a decrease in the number of cells that maintained transmembrane pH gradient. At the same time a decrease in the cell count on nonselective agar plates was observed.
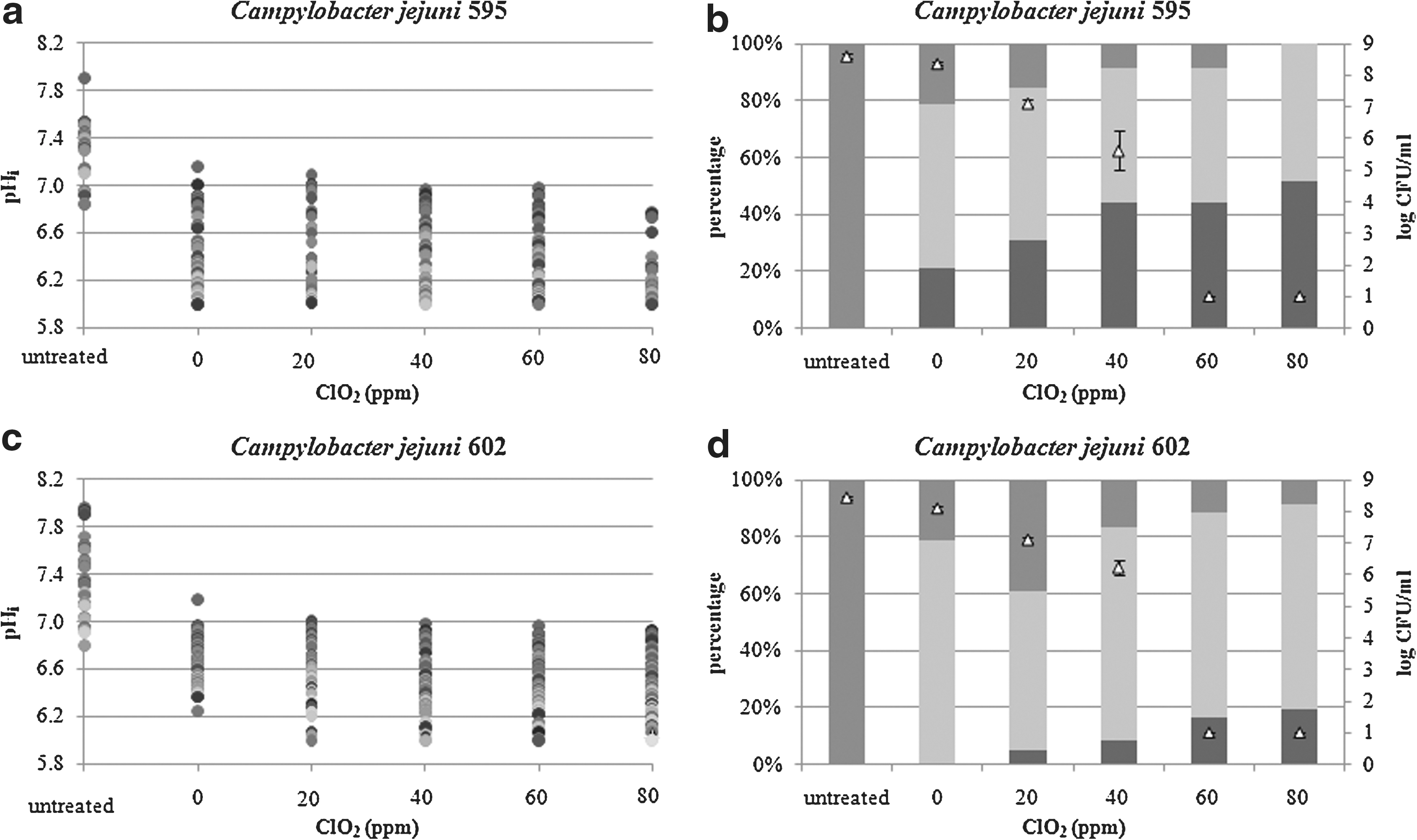
The intracellular pH (pHi) values of single cells
Our results revealed that a subpopulation of C. jejuni cells were able to maintain pHi > 6.8 when exposed to high ClO2 concentrations (60 and 80 ppm). This subpopulation present healthy cells as judged on the basis of their pHi (Rechinger and Siegumfeldt, 2002). However, the number of cells on nonselective agar plates was below the limit of detection (1 log CFU/mL). When the behavior of cells treated with 60 ppm for 2 min was further investigated under favorable growth conditions (Bolton broth pH 7.0, 37°C, microaerophilic atmosphere, data not shown), no growth was detected. The subpopulation of cells able to maintain pH gradient detected by the FRIM method was not culturable even under favorable growth conditions and may be dying.
Regarding the response of two C. jejuni strains when treated with different ClO2 concentrations, the obtained level of reduction was higher for C. jejuni 595 than for C. jejuni 602. At the same time their pHi values were also slightly different, with percentage of C. jejuni 595 cells not able to keep transmembrane gradient being greater than percentage obtained for C. jejuni 602.
In conclusion, our FRIM results obtained at the single cell level have shown that ClO2 treatment induces diversity among C. jejuni cells into subpopulations based on measured pHi. The heterogeneity in bacterial response to ClO2 stress would not be recognized if only average pHi values were obtained. Thus, it is very important to determine, where possible, physiological parameters of single cells. Increases in the concentration of ClO2 resulted in a higher proportion of cells with dissipated pH gradient and was accompanied by a decrease in number of culturable cells. Using FRIM, we determined subpopulation of the cells with positive gradient even though they lost their culturability. Whether the subpopulations of the C. jejuni cells found in vitro will occur in ClO2-treated food and what might be the effect of food matters on the occurrence and distribution of the subpopulation of the cells is not currently known. More research in this field is required since the presence of subpopulations of injured cells in food could present a potential risk for public health, especially for pathogens with low infectious dose. This emphasizes the need for incorporation of additional hurdles in food safety application to control the recovery and survival of injured cells. At the same time these results imply a need for further investigations of post-decontamination microbial behavior on the level of single cells, using also other tools that would be able to differentiate cells based on their viability and metabolic activity.
Footnotes
Acknowledgments
The research performed has been part of the project FOOD-CT-2005-007081 (PathogenCombat) supported by the European Commission through the Sixth Framework Program for Research and Development.
Disclosure Statement
No competing financial interests exist.