
Abstract
The objective of this study was to determine the frequency of nasal carriage of methicillin-resistant Staphylococcus aureus (MRSA) in slaughter pigs, to characterize the recovered isolates, and to investigate their genomic relatedness. Nasal swabs were collected from 53 finishing-pigs (F-pigs) and 53 suckling-piglets (S-piglets) at two different abattoirs in La Rioja (Northern Spain) coming from six production holdings. MRSA isolates were characterized by spa−, agr−, SCCmec−, and multilocus sequence typing, pulsed-field gel electrophoresis (PFGE)-ApaI, toxin gene profiling, antimicrobial susceptibility, and determination of antimicrobial resistance genes. MRSA isolates were recovered from 11 F-pigs (14 isolates) and 26 S-piglets (30 isolates). Forty of the 44 MRSA presented the spa-types t011, t108, t1197, and t2346, which corresponded to the sequence type ST398 and to the clonal complex CC398. Interestingly, the remaining four isolates from F-pigs presented the spa-type t3992, and they were ascribed to a new sequence type named ST1379 (a single-locus variant of ST97), which was included in clonal complex CC97. Five PFGE-ApaI clusters with up to nine individual patterns detected among our MRSA and low genomic relatedness was observed between F-pig and S-piglet isolates. All MRSA were positive for hla, hld, and hlg hemolysin genes. ST1379 isolates harbored eta, lukE/D, and hlg-2 toxin genes, whereas ST398 isolates were positive for hlb. A great variety of distinct resistance gene patterns were observed, most of them coming from F-pig isolates. MRSA virulence properties seem to be dependent of the isolate clonal lineage. This study showed that slaughter pigs are frequently colonized by MRSA CC398; moreover, the detection of strains belonging to CC97 underlines that other lineages are also able to spread in livestock. Further studies should assess the risk of CC398 and non-CC398 MRSA to enter the food chain as well as the human health implications.
Introduction
There is current concern about LA-MRSA in Spain, considering that this country accounts for a high rate of pig production holdings; nevertheless, data on the prevalence in pigs are still scarce (EFSA, 2009a, 2009b). The aim of this study was to investigate the carriage rates of MRSA in pigs of two distinct age groups at slaughter in Spain, to characterize the recovered isolates using distinct molecular techniques, and to determine the clonal diversity of the studied isolates.
Materials and Methods
Origin of samples
Individual nasal swabs were collected from 106 healthy pigs in La Rioja (Northern Spain) between September 2008 and March 2009. Samples corresponded to 53 finishing-pigs (F-pigs) and 53 suckling-piglets (S-piglets). Sampling was implemented on four different occasions (two for F-pigs and two for S-piglets) at two abattoirs, one for each age-group, with a slaughtering capacity of about 300 pigs per day. Tested animals came from six different farrow-to-finish holdings (Table 1).
Same holding of origin.
MRSA, methicillin-resistant Staphylococcus aureus; F-pigs, finishing-pigs; S-piglets, suckling-piglets.
Bacterial isolates and polymerase chain reaction screening of samples
Swabs were inoculated into Brain-Heart-Infusion broth supplemented with 6.5% NaCl and incubated at 37°C for 24 h; an aliquot of 100 μL was then streaked on Oxacillin-Resistant-Staphylococcal-Agar-Base (Oxoid) plates supplemented with 2 μg/mL of oxacillin (Oxoid) and incubated at 37°C for 24–48 h. Up to three blue presumptive MRSA colonies were selected from each positive sample and preserved for further studies. Identification of S. aureus was based on colony morphology, Gram staining, and catalase and DNase activities. MRSA identification was confirmed by detection of nuc (S. aureus specific) and mecA (methicillin-resistant staphylococci specific) genes by multiplex polymerase chain reaction (PCR), as previously recommended (CRL-AR, 2009).
PFGE of MRSA strains
Due to the non-typeability of ST398 strains using SmaI enzyme (caused by a restriction/methylation system leading to protection from SmaI digestion), strain relatedness was analyzed by PFGE of total DNA restricted with ApaI enzyme applying the HARMONY protocol guidelines (Murchan et al., 2003). The switching times of electrophoresis for ApaI digests were those implemented by Kadlec et al. (2009). PFGE patterns were analyzed using GelCompar software package (Applied Maths). To determine the percentage of similarity necessary for strain discrimination, reproducibility studies were conducted (López et al., 2008), analyzing by duplicate the macrorestriction patterns of 5 MRSA ST398/CC398 isolates. PFGE groups were defined by those formed at 89% cut-offs. The similarities between profiles were calculated using the Dice coefficient, with a maximum position tolerance of 1.2%, and the Unweighted Pair Group Method using Arithmetic averages.
Molecular typing of MRSA strains
All MRSA isolates were characterized by spa-typing, determination of staphylococcal cassette chromosome mec (SCCmec), and agr allotype by specific PCRs (
Toxin gene profiling
The presence of the genes encoding the Panton-Valentine-Leukocidin (PVL) toxin (lukS/F-PV), LukM-LukF-PV (lukM), and LukE-LukD (lukE/D) leukocidins were tested by PCR (Lina et al., 1999; Jarraud et al., 2002). Likewise, MRSA isolates were also screened by PCR for genetic determinants of hemolysins (hla, hlb, hld, hlg, and hlgv), exfoliative toxins (eta and etb), 18 enterotoxins (sea, seb, sec, sed, see, seg, seh, sei, sej, sek, sel, sem, sen, seo, sep, seq, ser, and seu), as well as for the toxic-shock syndrome 1 toxin (tst) (Jarraud et al., 2002; Hwang et al., 2007).
Antimicrobial susceptibility testing and detection of resistance genes
Antimicrobial susceptibility testing to 16 antimicrobial agents (Fig. 1) was performed by disk-diffusion agar (CLSI, 2009). Streptomycin, fusidic acid, and mupirocin breakpoints were considered as recommended by the Societé Française de Microbiologie (
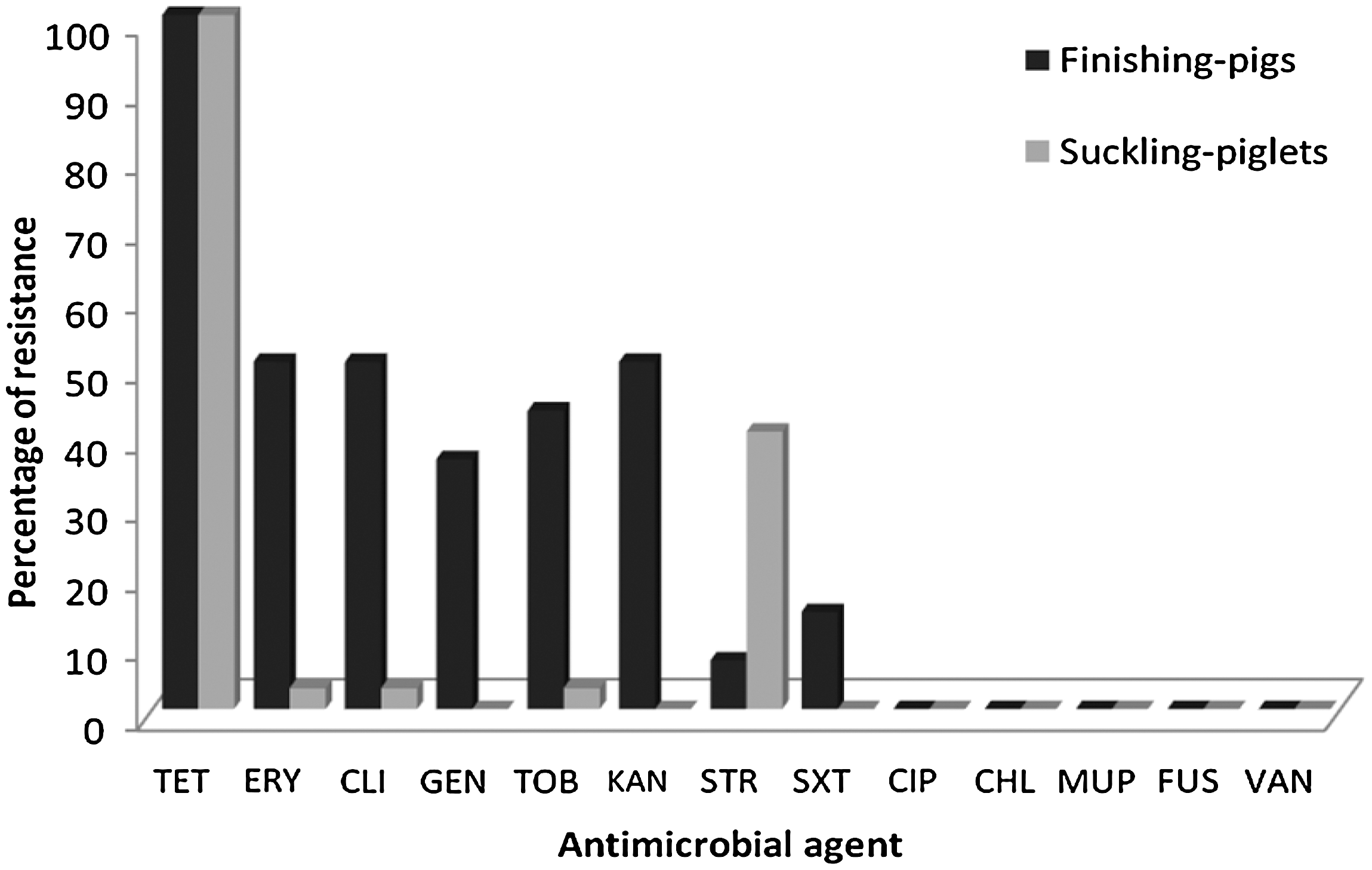
Comparison of non-beta-lactam antimicrobial resistance patterns of MRSA isolates coming from finishing-pigs versus those isolated from suckling-piglets. TET, tetracycline; ERY, erythromycin; CLI, clindamycin; GEN, gentamicin; TOB, tobramycin; KAN, kanamycin; STR, streptomycin; SXT, trimethoprim-sulphametoxazole; CIP, ciprofloxacin; CHL, chloramphenicol; MUP, mupirocin; FUS, fusidic acid; VAN, vancomycin; MRSA, methicillin-resistant Staphylococcus aureus.
GenBank accession number is indicated within brackets.
Results
Detection and isolation of MRSA
MRSA were detected in 11 of 53 F-pigs (21%) and in 26 of 53 S-piglets (49%) corresponding to 5 of 6 of the sampled production holdings. Forty-four MRSA isolates (14 of F-pigs and 30 of S-piglets) were recovered from the MRSA-positive animals and were further characterized (Table 1).
Molecular typing of studied strains
Genetic relatedness by PFGE after ApaI digestion of the chromosomal DNA of our MRSA strains yielded a relatively wide range of fragment profiles. Cluster analysis generated five major clusters (A–E), with nine individual patterns. The resulting dendrogram, its correlation with the other typing methods implemented, and the origin of the studied isolates are shown in Figure 2.
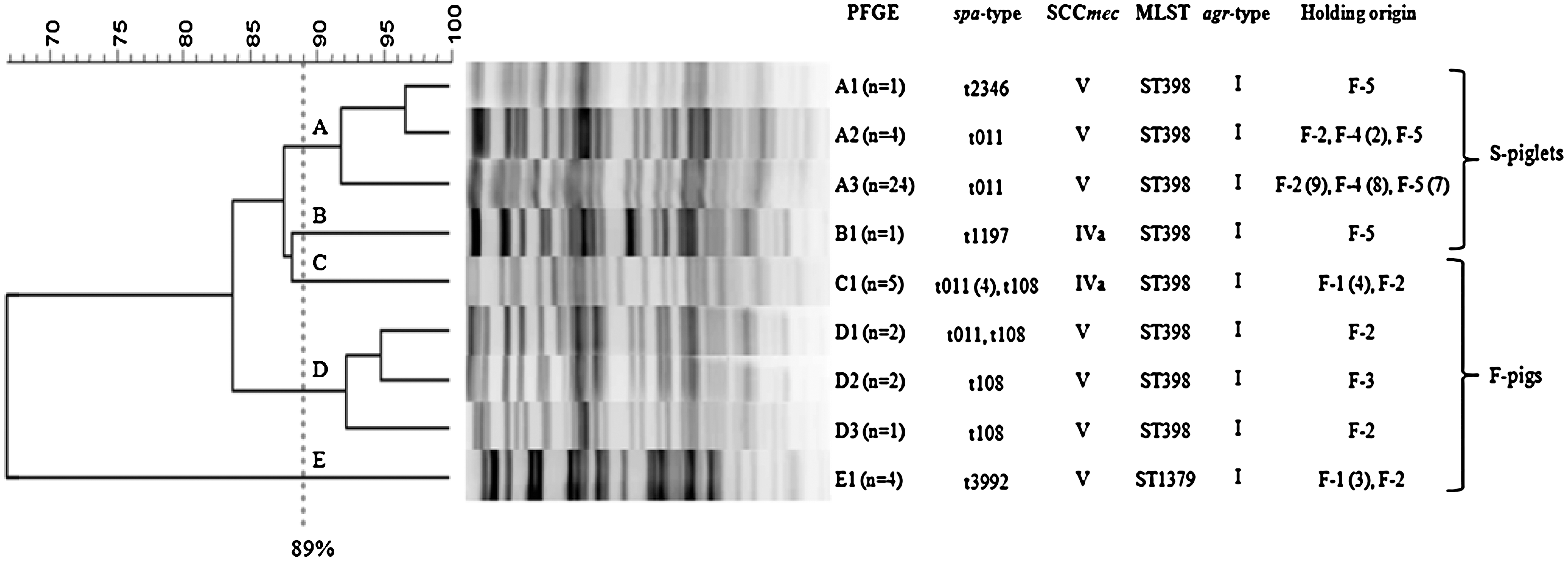
Dendrogram showing the relatedness between ApaI macrorestriction fragment profiles generated after PFGE from the MRSA ST398/CC398 and ST1379/CC97 isolates studied. A cut-off at 89% similarity was used to distinguish the five major clusters (labeled A to E), and 9 individual patterns were obtained (labeled A-E plus number). The spa-type, SCCmec cassette, MLST, agr-type, holdings of origin and animal type associated with each ApaI fingerprint are also indicated. The numbers in brackets after the different fragment profiles indicate the number of isolates that exhibited the respective fragment type. PFGE, pulsed-field gel electrophoresis; SCCmec, staphylococcal cassette chromosome mec; MLST, multilocus sequence typing.
Three spa-types were identified among the 14 MRSA isolates from F-pigs (number of isolates): t011 (5), t108 (5), and t3992 (4) (Table 3). The spa-types t011 and t108 are characteristic of the CC398 lineage (EFSA, 2009b; Vanderhaeghen et al., 2010), as was confirmed by MLST on six selected strains (one per PFGE type and spa-type) (Fig. 2). MLST analysis of the four isolates with spa-type t3992 revealed a new allelic profile, now classified as ST1379. This new ST is a single-locus-variant of ST97, presenting a single nucleotide mutation (C110A) at tpi locus, resulting in a nonsynonymous amino acid change (Ala37Asp). Consequently, ST1379 belongs to the clonal complex CC97. In this regard, 7 of 11 (63.6%) MRSA-positive F-pigs were colonized by CC398-MRSA, and the remaining ones (36.3%) carried MRSA non-CC398. MRSA recovered from F-pigs were typed as either SCCmecIVa (five isolates) or SCCmecV (nine isolates). SCCmecIVa was identified in four of five isolates spa-type t011 and in one of six isolates spa-type t108 (Table 3).
Number of isolates when more than one is included.
spa-type presented in the 4 ST1379/CC97 isolates.
TET, tetracycline; ERY, erythromycin; CLI, clindamycin; GEN, gentamicin; TOB, tobramycin; KAN, kanamycin; STR, streptomycin; SXT, trimethoprim-sulphametoxazole.
PFGE, pulsed-field gel electrophoresis; SCCmec, staphylococcal cassette chromosome mec.
Twenty-eight of the 30 MRSA isolates recovered from S-piglets presented the spa-type t011, one isolate was typed as t2346, and the remaining one was typed as t1197. These three spa-types are associated with CC398, as confirmed by MLST analysis of four selected strains. All isolates were associated with SCCmecV except for the MRSA t1197 strain, which harbored SCCmecIVa (Table 3). All 44 MRSA studied revealed agr type I.
Toxin gene profile and antimicrobial resistance pheno- and genotypes
All MRSA isolates were negative for the PVL toxin genes (lukS/F-PV) as well as for lukM, etb etd, tst, and all tested enterotoxin genes. All strains, independently of their clonal complex, were positive for hla, hld, and hlg genes. However, only ST1379/CC97 harbored the eta, lukE/D, and hlgv genes, whereas all ST398/CC398 isolates were positive for the hlb gene.
As for their antimicrobial resistance phenotypic characteristics, all MRSA strains showed resistance to beta-lactams and tetracycline, and different profiles were exhibited for the rest of the tested antimicrobial agents, which were dependent of the age-group (Fig. 1). Multi-resistance patterns (resistance to at least three classes of antimicrobial agents, in addition to beta-lactams) were exhibited in six MRSA strains from F-pig origin and only in one isolate from S-piglet origin.
MRSA isolates from F-pigs harbored more antimicrobial resistance genes than those of S-piglets and also exhibited a greater variability in the antimicrobial genotype profiles (Table 3). All 44 MRSA isolates carried the mecA and blaZ genes related to beta-lactam resistance. All isolates were resistant to tetracycline, and tet(K), tet(M), and/or tet(L) genes were present in different combinations. Among the eight macrolide-lincosamide-resistant strains, seven came from F-pigs and just one from S-piglets, and they harbored erm(A), erm(C), erm(T), and/or msr(A) genes in diverse combinations. All these isolates showed constitutive expression of resistance to macrolide-lincosamide-streptogramin B (MLSB), as shown by D-test. Gentamicin resistance was only detected in five isolates obtained from F-pigs, all of which carried the aacA/aphD gene. Kanamycin and tobramycin resistance was exhibited in six isolates from F-pigs, being mediated by the aadD gene (associated with aphA3 in one strain). Twelve MRSA strains (11 from S-piglets and 1 from F-pig) showed streptomycin resistance, and the str with/without aadA gene were identified. Two strains from F-pig origin were resistant to SXT, and both revealed the same combination of genes: dfrA + dfrG + dfrK.
Discussion
A high rate of MRSA detection among the production holdings tested was observed in this study. A recent European survey showed that the prevalence of breeding holdings positive for MRSA was 27%, and the prevalence found in Spain was 50% (EFSA, 2009b). However, that survey was conducted at farm level on dust samples, and MRSA prevalence might be different due to the sampling procedure.
Additionally, a high occurrence of MRSA carriers within slaughter pigs is detected in the present study (21% F-pigs, 49% S-piglets). Very few studies have so far pointed animal age as a possible factor that might influence LA-MRSA prevalence, which have obtained diverse results (Vanderhaeghen et al., 2010). Therefore, further investigations should try to unveil this possible influence of age, using a larger number of animals.
Most of our MRSA isolates belong to the CC398 (overall prevalence of 91%) with the remaining strains being ST1379/CC97. In the European study (EFSA, 2009b), 92% of MRSA were CC398, whereas 8% of strains were non-CC398, with ST1/CC1 and ST97/CC97 being the most frequently recovered. It is important to remark that in the European survey, MRSA CC97 was not detected among MRSA isolated from Spain.
The lineage CC97 is traditionally closely associated with cattle, and MRSA belonging to this clonal complex have been detected worldwide, being in most cases responsible for bovine mastitis (Smith et al., 2005; Aires de Sousa et al., 2007; Sung et al., 2008; EFSA, 2009b). Moreover, methicillin-susceptible S. aureus (MSSA) and MRSA CC97 have also been reported on human clinical isolates obtained in different countries (Feil et al., 2003; Chung et al., 2004; Ellington et al., 2008; Sung et al., 2008). In these human infections, no information on the possible animal contact was given. Additionally, the first description of CC97 in pigs was reported on MSSA, causing a swine infection in France (Armand-Lefevre et al., 2005), and recently the first detection of CC97-MRSA on clinical isolates in pigs in Germany has been published (Meemken et al., 2009) and from healthy slaughter pigs in Italy (Battisti et al., 2010). Further, a recent publication detected one CC97-MRSA on a rat living on a pig farm in Denmark (van de Giessen et al., 2009).
As far as we know, no contact between pigs and dairy cattle along the breeding or production chain occurred in our study. Moreover, CC97-MRSA strains came from two different farrow-to-finish holdings and were sampled at different periods. Due to the traditional close association of this clonal lineage with mastitis in cattle as well as its presence in human infections, epidemiological studies on the potential ways of inter-species transmission as well as investigations of the human health implications of the non-CC398 MRSA detected should be definitively addressed.
Data analysis of PFGE macrorestriction fragments showed remarkable clustering results given that groups seem to cluster according to their age group and in line with their SCCmec cassette. Several recent reports have shown a closer genomic background within CC398 isolates with the same SCCmec type (SCCmecIVa or SCCmecV) after digestion with Cfr9I enzyme, which also suggest that CC398-MSSA strains could have acquired the SCCmec cassette at different stages and then evolve and diversify (Argudín et al., 2010; van Wamel et al., 2010). This is the first study showing this association when ApaI enzyme was used. Further, this enzyme also allowed direct comparisons of CC398 and non-CC398 S. aureus isolates. It is interesting to remark that the same spa-types were present in different clusters and sub-clusters detected by PFGE. Our results point to the suggestion that distinct lineages within CC398 could be better adapted to different animal ages. In this sense, future investigations addressing this observation will be performed.
In our study, the multi-resistance genotype is generally associated with the presence of SCCmecIVa, differing from typical community-acquired-MRSA-SCCmecIVa (Vanderhaeghen et al., 2010). On the other hand, the only multi-resistant MRSA isolates carrying SCCmecV were two CC97 strains. There are only two studies reporting the SCCmec cassette on CC97 isolates, which detected SCCmecIVa and SCCmecV, respectively (Chung et al., 2004; Ellington et al., 2008). Likewise, this is the first report describing the antimicrobial genetic patterns of CC97 strains.
None of our isolates carried the PVL-encoding genes. PVL-positive CC398-MRSA isolates are uncommon; however, since its first finding from humans with no exposure to animal husbandry in China (Yu et al., 2008), its detection has been described on several occasions, always in isolates causing human disease (van Belkum et al., 2008; Welinder-Olsson et al., 2008). There is no report on PVL-positive CC398-MRSA isolated from livestock so far, differentiating these LA-MRSA strains obtained from animals from typical community-acquired-MRSA, which are characteristic for the presence of these toxin genes. On the other hand, PVL-positive CC97-MRSA isolates have not yet been reported.
All MRSA studied showed a low number of virulence determinants. Interestingly, the presence of eta and lukE/D genes as well as hlgv was homogeneous in the four CC97-MRSA isolates. LukED is closely associated with bovine mastitis caused by S. aureus and classically seldom detected in pigs (Yamada et al., 2005; Battisti et al., 2010), which emphasizes the possible bovine origin of these isolates. In addition, a great prevalence of these genes has been reported among clinical human S. aureus isolates (Vandenesch et al., 2003), which remarks the potentially higher pathogenic properties of our CC97 isolates comparing with those CC398 recovered.
As for the enterotoxin genes tested, very few data are available on the presence of these genes in CC398-MRSA strains of animal origin (Kadlec et al., 2009; Vanderhaeghen et al., 2010). Regarding CC97-MRSA isolates, a previous report showed the presence of few of these genes in ST97-MRSA obtained from clinical human isolates (Ellington et al., 2008). However, to the best of our knowledge, this is the first report that addresses the presence or absence of these genes in CC97-MRSA of animal origin. Although in our study the presence of these genes was negative, in-depth virulence studies should be conducted on these lineages to elucidate the potential enterotoxigenic properties of these strains, due to the potential impact on human health.
All MRSA strains studied in this report (either CC398 or CC97) showed resistance to tetracycline, which is a common trait among S. aureus of animal origin. The resistance gene tet(L) has been rarely detected in MRSA isolates (Kadlec et al., 2009). Hence, it is remarkable that 7 out of 14 strains from F-pigs harbored this gene. Alternatively, none of the MRSA strains from S-piglets carried tet(L) gene. This underlines that the phenotype of resistance seems to be associated not only to the animal age but also to the presence of particular genes.
Published data on MRSA from livestock have reported erythromycin-clindamycin resistance (Witte et al., 2007; Kadlec et al., 2009; Battisti et al., 2010; Mulders et al., 2010). Nevertheless, due to the absence of D-test analysis, there are no data on the prevalence of either constitutive or inducible resistance until now. Consequently, the constitutive expression of MLSB resistance revealed in our study is noteworthy, as it may be of special interest for treatment of infections caused by these organisms. It is interesting to remark that all MLSB resistant isolates harbored the erm(T) gene. This gene has been recently detected for the first time in staphylococci, being located in a plasmid (pKKS2187) also carrying tet(L) and dfrK genes (Kadlec and Schwarz, 2010). Nevertheless, in our isolates, these genes were not detected by PCR, suggesting either another mediator or a mechanism of recombination and subsequent loss of the plasmid. This finding will be addressed in future investigations. Further, this is the first description of erm(T) in non-CC398 S. aureus isolates.
As for the two SXT-resistant MRSA isolates, the aforementioned dfrK gene was identified, which also harbored tet(L) gene. This is in accordance with Kadlec and Schwarz (2009), who recently described this gene among porcine CC398-MRSA and demonstrated the physical linkage of both genes in a plasmid (pKKS25).
In addition, within the streptomycin resistance genes, and to the best of our knowledge, the str-positive isolates as well as the aadA gene have not been reported in CC398-MRSA strains earlier.
This report evidences a high capacity of CC398 lineage to acquire and maintain resistant determinants, given that for most resistance properties more than one resistance gene was present, especially in F-pigs. This observation is in line with CC97 resistance patterns detected, underlining that two of four strains were multi-resistant.
Conclusions
To our knowledge, this is the first comprehensive molecular characterized study on MRSA in pigs at slaughter in Spain and the first detection of CC97-MRSA in Spain. The resistance gene acquisition capacities of all isolates studied (CC398 and CC97) is of particular significance, because both clonal lineages can colonize various animal and human hosts. PFGE macrorestriction fragments also show the usefulness of PFGE-ApaI analysis for distribution of CC398-MRSA isolates into groups and subgroups as well as for direct comparison with non-CC398 strains. Epidemiological studies on the circulating lineages in diverse animal ecosystems should be conducted, to elucidate the transmission routes of MRSA lineages among animals and humans as well as their rates of colonization and infection. Similarly, age of animals must be considered when designing and comparing studies, to estimate the most relevant animal ages in terms of animal and human implications, on which comprehensive studies on MRSA should be addressed.
Footnotes
Acknowledgments
E. Gómez-Sanz has a fellowship from the Gobierno de La Rioja of Spain; and C. Lozano has a fellowship from the Ministerio de Ciencia e Innovación of Spain. We thank the veterinarians J. Somalo and A. Arana for their support during sample collection. This work was partially supported by Project SAF2009-08570 from the Ministry of Education and Science of Spain and FEDER.
Disclosure Statement
No competing financial interests exist.