
Abstract
Monoclonal antibodies (Mabs) specific for lipopolysaccharide (LPS) of Salmonella Enteritidis were evaluated in a model of LPS conjugated synthetic polymer immunization of Balb/c mice by conventional hybridoma method. Nine hybridoma cell lines were determined as antibody positive against LPS. The clone 5A8 secreting the highest antibody was selected for further characterization. Evaluated results indicate that the synthetic polymer can be used as an effective adjuvant in immunization with LPS, because the 5A8 Mab were obtained using synthetic polymer as an adjuvant. 5A8 Mab was classified as IgG2a isotype by antibody capture enzyme-linked immunosorbent assay. The reactivity of the Mab against lipid A and different LPS of Salmonella were investigated using an indirect enzyme-linked immunosorbent assay. Mab presented a wide spectrum of reactivity, coupling with antigens against Salmonella. The hybridoma 5A8 determined in this study has a great potential to be used in the development of diagnostic, prophylactic, and therapeutic agents specific for Salmonella Enteritidis and Salmonella LPS.
Introduction
S
Salmonellae are antigenically complex, and serovars have been differentiated by somatic lipopolysaccharide (LPS) or flagellar protein antigens. On this basis, they have been considered as possible candidates for vaccines and diagnostic reagents (Malik et al., 2002). Conventional methodology for identifying Salmonella is both labor intensive and time consuming, which requires several enrichment and selective platings followed by biochemical and serological identification. Salmonella identification can takes 7 to 11 days depending on the methods used (Mayer et al., 1985; Humbert et al., 1990; Keller et al., 1993; Adams and Moss, 1995; Solano et al., 2000; Schneid et al., 2005). Several more rapid serological assays to be used in screening Salmonella Enteritidis have been described; these include agglutination tests and enzyme-linked immunosorbent assay (ELISA) (Swaminathan et al., 1985; Wyatt et al., 1993) using different antigens such as LPS, flagellin, and so on (Kerr et al., 1992; Zijderveld et al., 1992; Solano et al., 2000). LPS is, therefore, a good antigen of choice for use in an immunochemical assay for the serological detection of antibodies (Luk and Lindberg, 1991; Jaoho et al., 2000; Brooks et al., 2008).
On the other hand, monoclonal antibodies (Mabs) have greater number potential for identifying, isolating, and purifying bacterial antigens and also in defining the role of specific antigens in diagnosis, classification, and immunity to infection. They may also facilitate serotyping and strain differentiation (Torensma et al., 1992; Malik et al., 2002). Development of the novel reagents such as diagnostics Mabs has become economically important for detection of Salmonella Enteritidis.
The objective of this study was the production of Mab against Salmonella Enteritidis LPS via the immunization of Balb/c mice with LPS conjugated synthetic polymer instead of whole bacteria and determination of serological specificities of the obtained Mabs for their potential use in developing novel reagents.
Materials and Methods
Bacterial strains
Salmonella Enteritidis 64K (1,9,12: g,m) strain, which was used for extracting LPS antigen, was kindly provided by Dr. K. Tayfun Carli from Uludag University, Faculty of Veterinary Medicine, Bursa, Turkey. Salmonella Typhimurium (1,4,5,12: -) and Salmonella Senftenberg (1,3,19: g,(s),t) strains that were used for the preparation of LPS antigen were kindly provided by Dr. Radka Ivanova from National Center of Infectious and Parasitic Diseases, Sofia, Bulgaria. Salmonella Typhimurium, Salmonella Senftenberg were used for the isolation of LPS in the confirmation studies of the isolated antigens. Cultures were maintained by three replicating subcultures through a single colony to enhance the purity, identity, and homogeneity of the culture. After that, streaked single colony checked by Gram stain technique was used in stock culture preparation. Salmonella Enteritidis bacteria grown in Brain Heart Infusion broth (Becton Dickinson) in bioreactor was used for the LPS isolation as described by Nalbantsoy et al. (2007).
Extraction of LPS
The LPS isolation was performed by modified Westpal procedure (Toman and Skultety, 1996). The lyophilized 1 g Salmonella cells were solubilized in 50 mM Tris-HCl buffer (100 mL, pH 7.5) and treated with 0.2 g MgCl2, 6H2O, 0.5 mg bovine serum albumin (Sigma), 100 μL RNase (100 μg/mL, EC 3.1.27.5), and 100 μL DNase (100 μg/mL, EC 3.1.21.1) both from bovine pancreas (Boehringer) shaking overnight at 37°C; then enzymes were inactivated for 10 min at 75°C; and then treated with 100 μL trypsin (300 μg/mL EC 3.4.21.4, from bovine pancreas; Serva) for 90 min at 37°C. Enzyme inactivation has been repeated as just described; and 100 μL proteinase K (EC 3.4.21.14, from Tritirachium album; Sigma) was added and incubated shaking overnight at 37°C. Enzyme inactivation has been repeated, and the suspension was centrifuged at 30,000 × g for 1 h. After the pellet was incubated in 100 mL acetone at room temperature (RT), the cell suspension was centrifuged at 1500 × g for 15 min. The cells were further extracted with chloroform-methanol (Sigma), (2:1, v/v) at 20°C overnight. The cell suspension was centrifuged at 1500 × g for 20 min. Then cells were suspended in preheated distilled water (50 mL, 68°C) and extracted with an equal volume of aqueous 90% phenol for 20 min at 68°C by shaking. The extracts were cooled on ice, and the aqueous phase was collected after centrifugation at 1500 × g for 20 min. The phenol phase was re-extracted with water twice, and the combined aqueous phases were evaporated and dialyzed against distilled water for 3 days and freeze-dried.
Gel electrophoresis for LPS
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed in slabs containing 18% of polyacrylamide, and the gels were silver-stained for LPS as described by Toman and Skultety (1992). About 5 μg of LPS was loaded to the gel. Marker was purchased from Sigma.
Analysis of LPS
Estimation of 3-deoxy-D-manno-2-octulosonic acid (Kdo) residues was performed by a modified thiobarbiuric acid assay, and protein content was estimated by a modified Lowry assay as reported by Toman and Skultety (1996). The LPS (500 μg) was hydrolyzed in 0.1 mL 2 M trifluoroacetic acid (Sigma-Aldrich) (0.1 mL) at 100°C for 2 h. The residue was dissolved in 0.1 mL 1M ammonium hydroxide (Sigma-Aldrich) solution containing 1 mg sodium borodeuteride and incubated at RT overnight. The dried residues were acetylated with acetic anhydride (50 μL)-pyridine (50 μL) mixture at 100°C for 1 h. Then water was added and solvents were removed by evaporation. The residues were extracted with chloroform-water (Merck) (140 μL, 1:1, v/v) thrice; the organic layers were pooled, dried (Na2SO4), and evaporated. The acetylated samples were finally dissolved in chloroform and immediately analyzed as follows: Gas chromatography (GC). GC-mass spectrometry of aditol acetates was performed on a Finnigan MAT SSQ 710 mass spectrometer using a fused-silica capillary column SP-2330 (30 m × 0.25 mm; Supelco). The column temperature programs were 80°C (2 min) to 245°C at 30°C/min with a 36 min hold at a helium gas flow rate of 25 cm. s−1. Electron impact mass spectra were recorded at electron energy of 70 eV and an ion source temperature of 150°C.
Preparation of lipid A
Lipid A was released from LPS prepared by mild hydrolysis in aqueous acetic acid as described by Hussein et al. (2001). Briefly, the 0.5% concentration of LPS dissolved in 2% acetic acid was incubated at 100°C until the lipid A precipitated; the suspension was centrifuged at 14,000 × g for 20 min to pellet lipid A, and the pellet was washed thrice with ultra pure water and then freeze-dried.
Enzyme-linked immunosorbent assay
Indirect ELISA was used for confirmation of purified antigens. Briefly, 96-well polystyrene plates (Nunc) were coated with purified LPS (2 μg per well diluted in 100 μL 0.05 M carbonate/bicarbonate buffer, pH 9.6). After overnight incubation at 37°C, the plates were washed thrice. The diluted polyclonal and Mab supernatants specific for O:9 (produced by Dr. Ivanova) were added to coated wells with LPS, and the plates were incubated at 37°C for 1 h. After washing, secondary anti mouse IgG peroxidase (Sigma) and anti rabbit IgG peroxidase (Sigma) conjugates were added and incubated for 1 h at 37°C. The enzyme reaction was developed with the addition of H2O2 (Merck) and O-phenylenediamine (Sigma) and stopped after 30 min incubation by the addition of 4M H2SO4 (Riedel-de Haën) per well. The optical absorbance was measured at 492 nm.
Preparation of LPS conjugates with synthetic polymers
Polyelectrolyte (PE) complexes of LPS (negatively charged) were obtained from oppositely charged linear PEs. Polyacrylic acid (PAA)-Cu2+ and two modified different poly-4-vinylpyridine (P4VP), P4VP-ethyl bromide (P4VPEtBr), and P4VP-ethyl sethyl bromide (P4VPEtStBr) were used in preparation of conjugates. The PVP polymer that is synthesized by Prof. Akdeste was modified in his laboratory (Yıldız Technical University, Department of Bioengineering, Istanbul, Turkey). PAA (100,000 MW) was commercially purchased from Aldrich. Modified P4VP compositions were analyzed by Fourier transform infrared (FTIR) spectrophotometer (Perkin-Elmer [ATR sampling] Spectrum One). The stable LPS-polymer complexes are approximately formed at pH 7.00. After antigen polymer complex preparation, conjugates were analyzed by Gel Permission Chromatography (Viscotek, TDA 302) and Zetasizer (Nano series, ZEN 3600; Malvern). To prepare a LPS-polymer complex, different amount of P4VPEtBr or P4VPEtBr and PAA were dissolved in phosphate-buffered saline (PBS) and then mixed with different concentrations of LPS dissolved in PBS. Stable LPS-Cu2+-PAA complexes were obtained by 60 μL 55 mM CuSO4.5 H2O (Merck) addition. After centrifugation at 3500 × g for 10 min, the supernatant was investigated for the presence of conjugation. LPS-Cu2+-PAA was analyzed only with Zetasizer for differentiation of the dimension.
Animals
Balb/c mice (5–6 weeks-old) were obtained from The Scientific and Technological Research Council of Turkey, Marmara Research Center (TUBITAK-MAM). Mice were maintained in groups of 10 under standard conditions of temperature 22°C ± 1°C with regular 12 h light: 12 h dark cycle and allowed free access to standard laboratory food and water. The experimental protocol was approved by the Local Ethics Review Committee for Animal Experimentation of Ege University (number 2007-36).
Determination of LD50
Animals weighing 20–25 g were used (n = 10 for each group). The number of animals that died during this period was expressed as percentage, and LD50 was determined by a Probit test using 50% death within 24, 48, and 72 h after intraperitoneal (ip) administration of ultra pure water (control group) and LPS at different doses (100, 200, 400, 600, and 800 μg/mouse).
Immunization
Balb/c (6–8 week-old) mice were immunized ip with the estimated ¼ of LD50 amount of LPS conjugates. The mice were boosted on day 21, 42, 64, 86, 108, 123, and 137 with 3/4 and then ½ amount of the initial injection amount. Antibody levels were checked on day 112 by ELISA. Finally, sera were obtained after terminal heart puncture on day 140.
Preparation of hybridoma cell lines, screening and characterization of MAbs
The mouse that developed the highest antibody response was sacrificed and its spleen was removed for use in the fusion process. Spleen cells were fused with nonimmunoglobulin producing × 63-Ag8-653 murine myeloma cells purchased from the HUKUK (Animal Cell Culture Collections) in Foot-and-Mouth Disease Institute (Ankara) of Ministry of Agriculture and Rural Affairs of Turkey. Briefly, spleen cells from the immune Balb/c mouse and myeloma cells were fused at a ratio of 10:1 in serum-free medium by using polyethylene glycol (molecular weight, 4000; Sigma) as the fusion agent. Hybridoma selection was carried out using Dulbecco's modified Eagle's medium Ham's F:12 (Biochrome) with hypoxanthine, aminopterin, and thymidine (Sigma) with nonimmune mouse spleen cells as feeders. After 10 to 14 days, supernatants were screened using a spot ELISA to detect hybridomas secreting specific antibodies for LPS of Salmonella Enteritidis. The selected hybridomas were cloned twice by limiting dilution and expanded for storage in liquid nitrogen after sterility check by conventional methods for microbiological contamination, 4′,6-diamidino-2-phenylindole (DAPI) (Sigma) staining, and administration of Gibco-mycotect (Gibco) kit for Mycoplasma contamination. Isotyping of Mabs was carried out with ELISA that used specific antibody class or subclass (Sigma) and with Western blot. The specificity of each MAb was determined in an indirect ELISA by testing each Mab against purified LPS from Salmonella Tymurium and Salmonella Seftenberg and purified lipid A from Salmonella Enteritidis. Chromosome slides were prepared by conventional techniques and stained by Giemsa (Cotton et al., 1980; Suomalainen et al., 1983; Kalantarova et al., 2002). Preparations of metaphase chromosome of myeloma and hybridoma cells were performed using standard procedures (Hasty et al., 1982; Herr et al., 1985; Kalantarova et al., 2002). Metaphase chromosome preparations were obtained by adding colchicines to cells during the exponential growth phase at 1.5–2 h. Cells were suspended in 0.075 M KCI (Merck) for 11–12 min, centrifuged, and fixed in glacial acetic acid or methanol (Merck; 1:3, v/v) for 20 min. Drops of the cell suspension were placed onto a cold, wet slide, dried, and stained with Giemsa (Sigma). Chromosome numbers were determined after analyzing a minimum 50 metaphase figures for each line.
Western blotting
SDS-PAGE and western blots were performed under standard conditions using hybridoma supernatant and LPS. Five, 15, 20 μL of hybridoma supernatant and 5 μg of LPS were loaded to the gel. Hybridoma supernatant or LPS was separated on 10%–12% gels and transferred to a polyvinylidene fluoride (PVDF) membrane (Amersham) by wet transfer blotter. PVDF membrane was blocked with 5% dry milk in Tris-Borate-Saline solution containing 0.1% Tween 20. Primary antibody was used for hybridoma supernatant; however, hybridoma supernatant was used as the primary antibody for LPS, and secondary antibody incubations were carried out by using Tris-Borate-Saline solution containing 0.1% Tween 20 containing 0.5% dry milk at RT for 1 h. Membranes were developed using 2 mL ECL Plus reagent (Amersham) for 5 min and were photographed by Kodak X-Ray (Kodak) films in a dark room.
Results
Isolation of LPS, lipid A, and LD50 of LPS
The LPS was isolated from the purified cells by a conventional hot phenol-water procedure in a yield of 6%. After dialysis and lyophilization, white opaque LPS was obtained as described by Lounatmaa (1985). The isolated LPS protein content was calculated as 3.6%. The LPS gave a single band (14 kDA) characteristic for the semi smooth LPS on SDS-PAGE (Fig. 1). Quantity of Kdo residues in LPS determined by thiobarbiuric acid assay was 83 nmol/mg. GC-mass spectrometry analyses of neutral sugar content of purified LPS and indirect ELISA with LPS specific Mab and Pab results are presented in Tables 1 and 2, respectively. Moreover, the indirect ELISA with LPS specific Mab and Pab results were given in Table 2. Lipid A was isolated from LPS in a yield of 3%, and it was used to identify the cross reactivity with Mab. Since LPS is an endotoxin, LD50 value was determined in Balb/c mice before application of the immunization procedure. For this purpose, the different doses of LPS were injected, and ip LPS quantity that kills 50% of the animals at 24, 48, and 72 h was determined. After administration of LPS, the animals exhibited symptoms specific for endotoxic activities, such as rapid breathing, fever, trembling, diarrhea, shock, and death (Heppner and Weiss, 1965; Kaufmann et al., 2001). The calculated value of LD50 was 450 μg/mouse at 72 h. The estimated approximate ¼ of LD50 concentration was used for immunization of the mice (100 μg/mouse LPS with polymer conjugates).
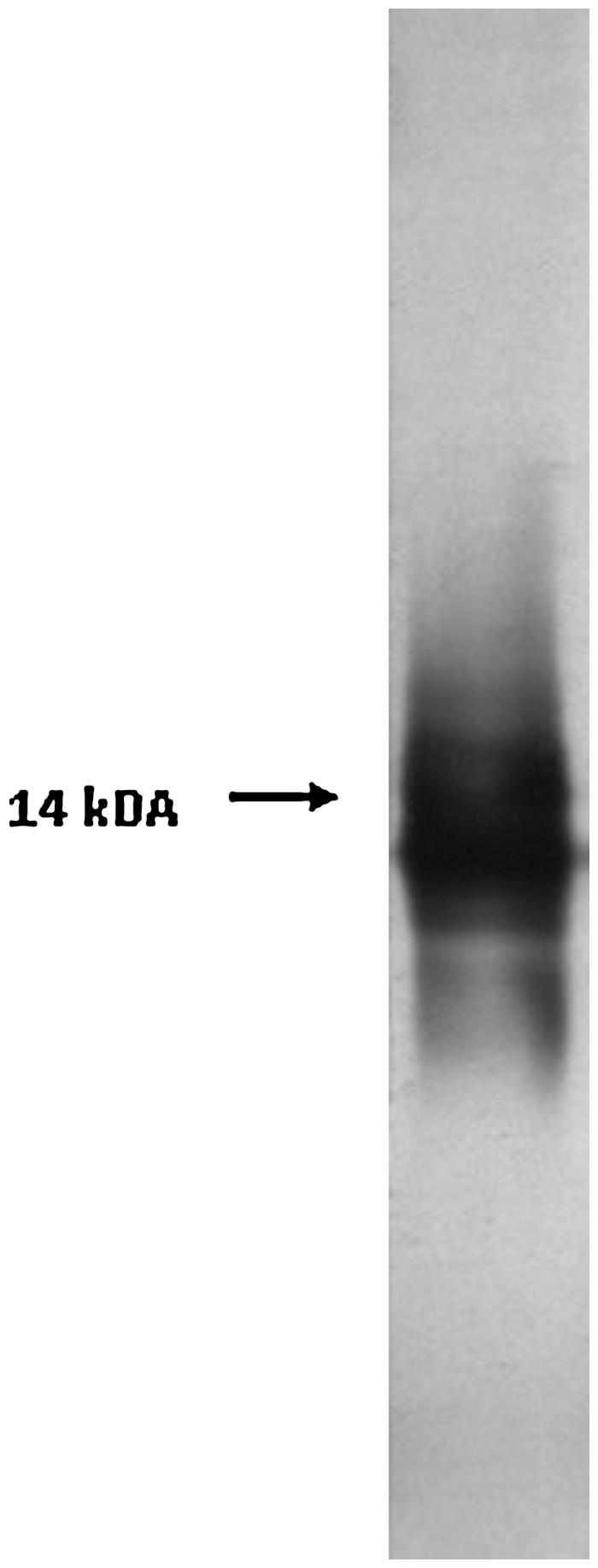
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis silver stains of the 5 μg LPS. LPS, lipopolysaccharide.
LPS, lipopolysaccharide.
O.D.492nm.
O:9 specific rabbit polyclonal antibody provided from NCIPD, Bulgaria.
O:9 specific Mab supernatants provided from NCIPD, Bulgaria.
ELISA, enzyme-linked immunosorbent assay; Mab, monoclonal antibody.
LPS conjugates with synthetic polymers and immunization
The modified P4VPEtBr and P4VPEtStBr from P4VP were controlled by FTIR spectrum with the comparison of the spectrum of P4VP. In addition, solubility of P4VPEtBr and P4VPEtStBr in water was compared with P4VP, which is determined as water insoluble to identify their suitability for conjugate preparation. Results indicated that modification of the polymers was successfully performed. Later, LPS-polymer conjugates were prepared with varying amounts in PBS at ∼pH 7.0. The LPS-Cu2+-PAA complex was carried out in soluble form unlike insoluble LPS-P4VPEtBr and LPS-P4VPEtStBr complex. After preparation of conjugates as just mentioned, each conjugation was detected and compared for the differences in dimension and other properties (by Gel Permission Chromatography) from the origins; LPS, polymer and LPS-polymer complex individually as given in Figures 2 and 3, respectively. The experiments with different types of polymer have shown that the optimum conjugation ratio was 1:1 mg/mL−1 LPS-polymer for preparation of the suitable complex with all types of polymers. Then the conjugates and naked LPS were injected into 5–6 week-old Balb/c mice by ip at a concentration of 100 μg/mouse. The polyclonal antibodies from Balb/c sera rose against purified LPS. The antibody level in sera was found with antigen mediated ELISA between 128,500 and 179,500 on day 112 for naked LPS and varied from 37,900 to 126,100 for LPS-polymer that conjugates after the immunization of one prime and seven boosters. The results demonstrated the immunoreactive properties with LPS as IgG response. After administration of two more injections with naked LPS, the antibody level was found to vary from 86,700 to 283,300 at O.D.492 on day 142 for LPS polymer complex.

Analysis of LPS and PAA conjugation with Zetasizer. PAA, polyacrylic acid.
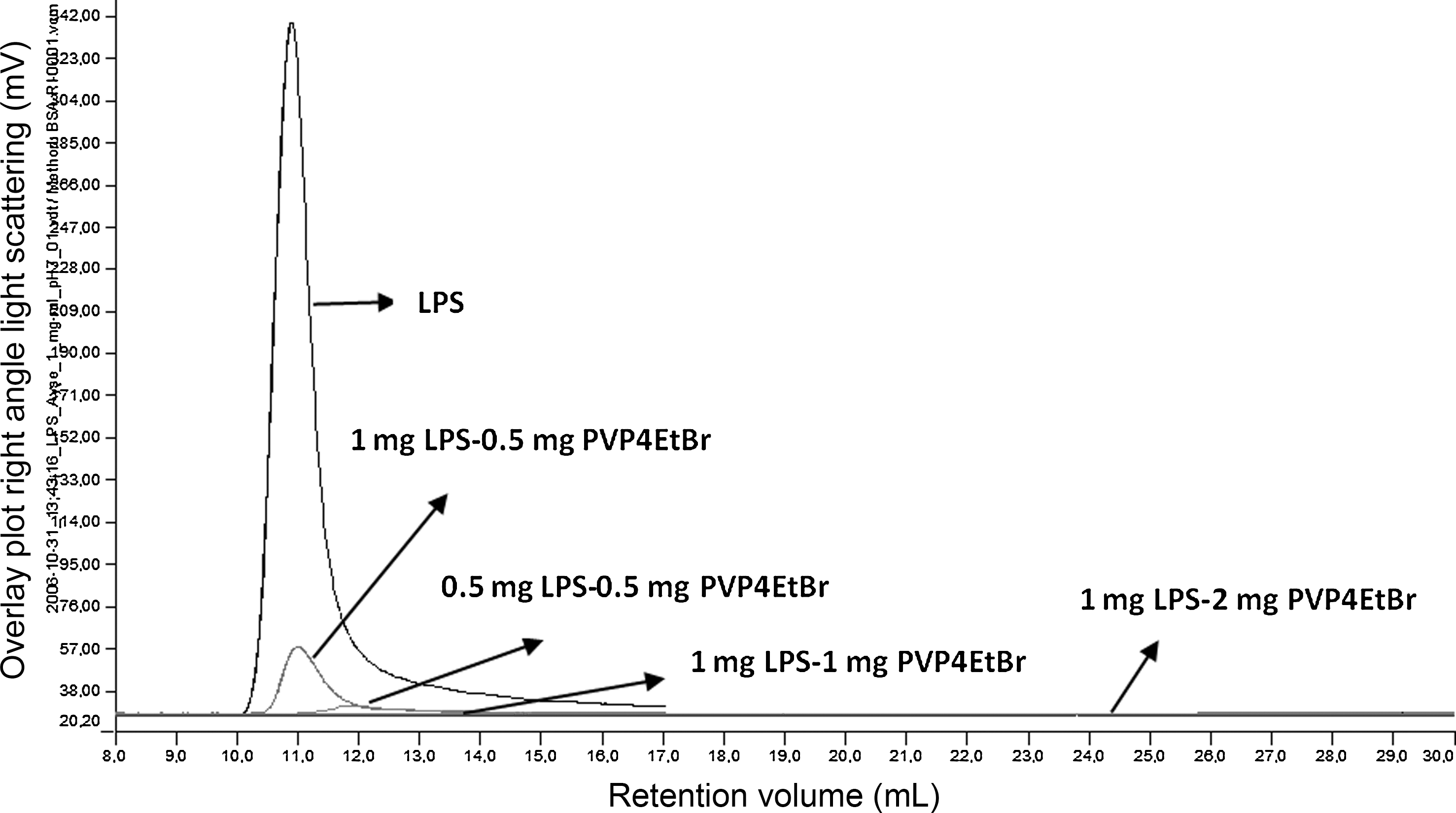
Analysis of LPS and PVP4EtBr conjugation by Gel Permission Chromatography (y-axis, overlay plot right angle light scattering [mV], x-axis, retention volume [mL]).
LPS-reactive hybridomas, screening and characterization
At the end of the immunization, fusion procedure was performed to mouse with the highest antibody titer. Fusing yielded a total of 97 hybridomas. After 10 to 14 days, supernatants were screened using an indirect ELISA to detect hybridomas secreting specific antibodies against LPS of Salmonella Enteritidis. Screening procedure resulted in five hybrid clones (in five fusions) reactive against the LPS antigens. Finally, one hybridoma was selected for the production of Mab after subcloning. The selected hybridoma was cloned twice by limiting dilution, and nine hybridoma cell lines were determined antibody positive against LPS. The clone 5A8 that secreted the highest antibody concentration was selected for further characterization. Isotyping results classified Mab as the subclass IgG2a by antibody capture ELISA. The reactivity of Mab with different strains of Salmonella LPS was investigated using an indirect ELISA as shown in Table 3. Moreover, the Mab demonstrated reactivity with lipid A isolated from Salmonella Enteritidis (Table 4) (Mansfield and Forsythe, 2001). The Western blot analysis with hybridoma supernatant gave a double band, which is characteristic of the IgG antibodies specific for light chain (25 kDA) and heavy (55 kDA) chain as shown in Figure 4. Immunoblot analysis of the LPS with 5A8 hybridoma supernatant showed that the 5A8 Mab bound well to the LPS (Fig. 5). The mouse spleen cells contain 40 chromosomes (Sawada et al., 1983); estimated metaphase chromosome contents of the parental × 63-Ag8-653 myeloma cells and the antibody-secreting hybridoma cells were 43–68 and 63–88, respectively.
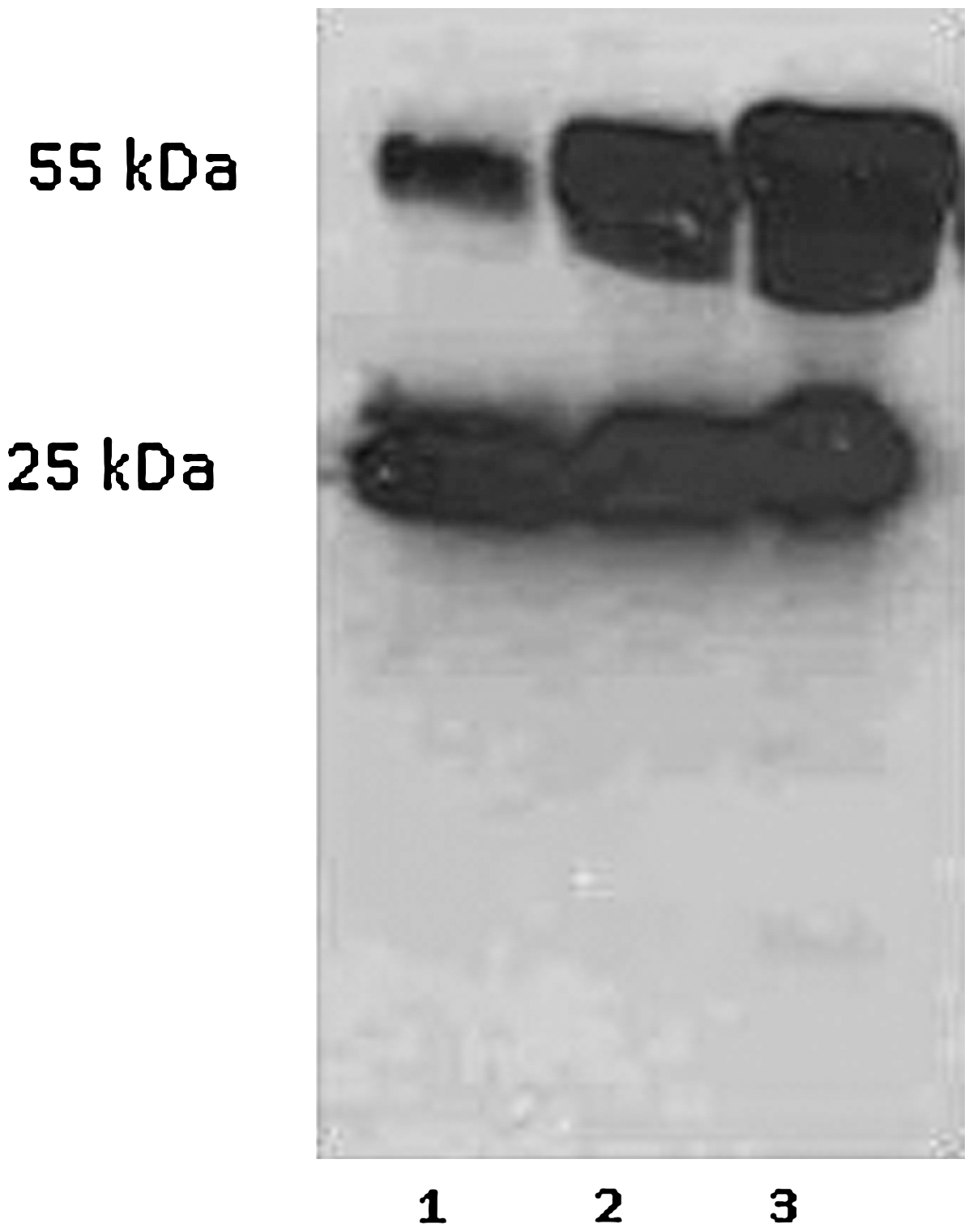
Western blot analyses of hybridoma supernatant with different amount (5, 15, and 20 μL).
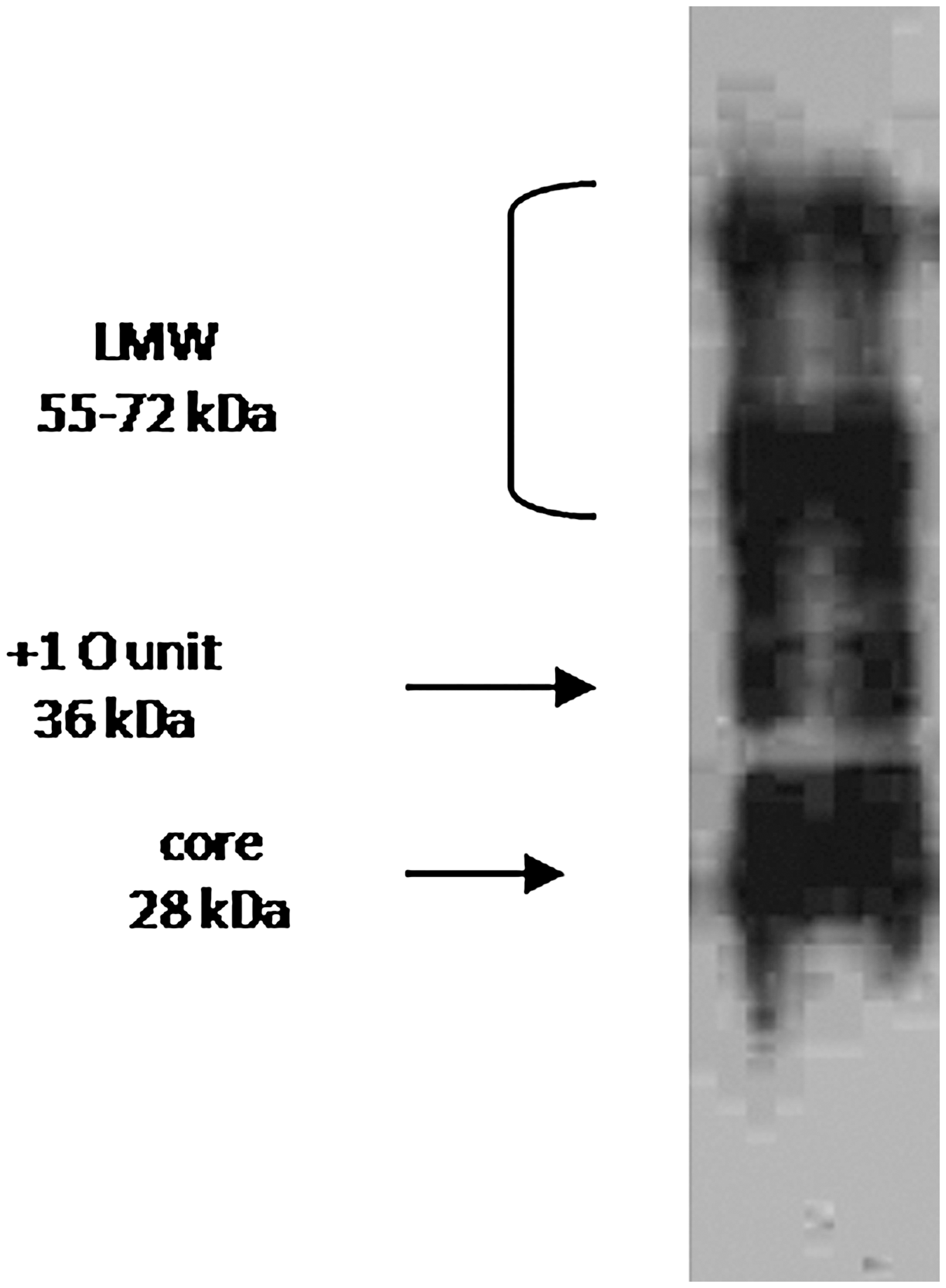
Immunoblot analysis of the purified LPS stained with hybridoma supernatant of 5A8.
O.D.492nm.
Culture supernatants.
O.D.492nm.
Culture supernatants.
Mouse anti sera specific for isolated LPS.
Discussion
In this report, the LPS specific Mab against the Salmonella Enteritidis was evaluated, which could be prospectively applied as diagnostic tools. In attempts to improve effective immunization strategy, we considered that immunizing animals with LPS mediated PE conjugates which increased amount of LPS antigens in a better form of presentation to the host for immune recognition.
We preferably focused on the use of purified LPS as immunogens. As expected, the purified LPS protein contamination was found typically between 1% and 3% per mg LPS by hot-phenol-water procedure (Lounatmaa, 1985). The SDS-PAGE profile of the LPS was low molecular weight (LMW), which was the characteristic of the repeated few sugar units (Rahman et al., 1997; Peter et al., 1999). The neutral sugar contents of purified LPS, rhamnose, mannose, galactose, and glucose, were found comparable to the previous studies (Nikaido, 1962; Lüderitz et al., 1971; Hitchcock and Brown, 1983; Steinbacher et al., 1996; Rahman et al., 1997). The presence of O-chain sugars rhamnose, mannose, galactose, and glucose (Peter et al., 1999) were identified with the isolated LPS that contained O chain. Ribose sugar residues were slightly contaminated with nucleic acid-related proteins. The immunodominant sugar tyvelose residue, which is the serovar designation for group D1 Salmonella, was not detected in the LPS examined in this study on the derivatization by GC method as reported by Peter et al. (1999). Moreover, the ELISA results (Table 2) indicated higher antibody levels than the results of Lüderitz et al. (1971) that related it to the semi smooth LPS containing repeated few sugar units (Peter et al., 1995). Semi smooth LPS were pathogenic unlike rough LPS or nonpathogenic strains (Peter et al., 1999). In this study, the serological evidence, based on using Salmonella Enteritidis O:9 specific Pab and Mab, indicated that the isolated LPS was in semi smooth structure.
The complexes or conjugate of nonimmunogenic synthetic PE with microbial and viral protein and polysaccharide antigens that were given before inoculation confer protection against diseases. The PE provides effective immune protection without traditional classical adjuvants as previously described (Petrov et al., 1990; Mustafaev, 2004). Another important point is that PE is nontoxic, biodegradable, and has an LMW and it is safe to use as an adjuvant (Mustafaev, 2004). In addition, LPS is a well-known endotoxic agent for the Balb/c mice; the death of none of the mice was determined after the administration at ¼ of estimated LD50 of LPS concentration. However, the mice exhibited slight symptoms related to the endotoxicity of LPS. Preliminary experiments indicated that the immunization with LPS protection in mice against experimental infections with bacteria possessing homologous with LPS in immune response and LPS could induce specific antibodies, generated a strong IgG response (Svenson et al., 1976; Svenson et al., 1979; Kenne and Lindberg, 1983; Mayer et al., 1985; Malik et al., 2002; Ochoa-Reparaz et al., 2005). At the end of immunization, the sera antibody titers were showed maximum value at 201,900 for naked LPS. On the other hand, the highest antibody titer against LPS obtained with PAA after the P4VPEtStBr using as adjuvants. It was found out that P4VPEtStBr was a much more effective adjuvant than the others. The 5A8 Mab showed to be weak to moderate reactivity against immunodeterminants of Salmonella LPS but a stronger degree of binding to LPS of Salmonella Enteritidis. It is more likely that cross-reactions are due to common epitopes present on different LPS. As pointed out earlier, the LPS represent the major surface antigens of a great variety of Gram-negative bacteria. Antisera to antigenic determinants in the R core and lipid A regions of the LPS molecule are able to protect experimental animals against lethal infection with Gram-negative bacteria and against a number of different pathophysiological effects induced by homologous and heterologous LPS (Ward et al., 1988). In particular, the core oligosaccharide exhibits distinct immunoreactive properties and is of great interest, because it may broadly induce cross-reactive antibodies (Ward et al., 1988; Mitov and Terziiski, 1991; Torensma et al., 1992; Mitov et al., 1993; Brooks et al., 2008). Therefore, it is suggested that the 5A8 hybridoma Mab is potentially useful for the development of improved diagnostic tests for the detection serogroup D1 Salmonella. Several authors mentioned that the findings also demonstrated the existence of similar or identical epitopes in the LPS antigens of Salmonella (Ward et al., 1988; Luk and Lindberg, 1991; Torensma et al., 1992; Zijderveld et al., 1992; Keller et al., 1993; Singh et al., 1999; Iankov et al., 2002, 2004; Malik et al., 2002; Mitov et al., 2003; Schneid et al., 2005). The presence of O-chain on the LMW LPS was supported by a Western blot analysis using hybridoma supernatant of 5A8, which is specific for the terminally linked tyvelose residue in the O-chain (Rahman et al., 1997; Peter et al., 1999). The analysis of metaphase chromosomes results that the indications were such that spleen cell chromosomes and myeloma chromosomes were rearranged to produce antibody as myeloma-spleen cell hybrid.
Conclusions
Salmonella Enteritidis is one of the major causes of food borne infections and most frequently isolated serovar in human salmonellosis in Europe and Turkey. Poultry-related food products are the most important sources for human Salmonella Enteritidis infection. In this report, we describe the 5A8 Mab as the subclass IgG2a showed weak to moderate reactivity against immunodeterminants of Salmonella LPS but strong degree of binding to LPS and lipid A of Salmonella Enteritidis. Isolated LPS were used with synthetic polymer as an adjuvant to immunization of the mice. 5A8 Mab was obtained using the adjuvant of P4VPEtStBr, which was considered as an effective adjuvant. In addition, P4VPEtBr and PAA are good candidates for use of adjuvant in immunization with LPS. Besides the 5A8 Mab that exhibited a broad cross-reactive action with endotoxin from a wide range of Salmonella LPS, the hybridoma 5A8 demonstrated in this study has also a great potential for use in the development of diagnostic, prophylactic, and therapeutic agents specific to Salmonella Enteritidis and Salmonella LPS.
Footnotes
Acknowledgments
We thank to Dr. Rudolf Toman from Slovak Academy of Sciences (Bratislava, Slovak Republic) for the technical advice and his help with the isolation and confirmation of LPS in his laboratory. We are grateful to the deceased Prof. Mustafa Akdeste and Assoc. Prof. Zeynep Akdeste for their technical and scientific support to obtain the LPS-synthetic polymer conjugates in their laboratory (Yıldız Technical University, Department of Bioengineering). Also, we thank Lokman Varıslı, M.sc. and Nuray Bogurucu for their kind assistance with the Western blot designation.
This work was financially supported in part by a grant from Research Fund of Ege University (2002FEN09).
Disclosure Statement
No competing financial interests exist.