
Abstract
We investigated the efficacy of bacteriophage-based detection technology to detect Escherichia coli O157:H7 from ground beef. The assay involved a short enrichment period of 8 h followed by capture of the pathogen on O157-specific immunomagnetic beads. The captured cells were treated with O157-specific lytic bacteriophage, CSLO157. Upon phage-induced lysis, the enzyme adenylate kinase, which was released from the lysed cells, was measured in terms of relative light units using luciferin–luciferase assay. The plaque forming efficiency (e.g., phage susceptibility) and ability to capture cells with immunomagnetic beads were examined using an array of 74 E. coli O157:H7 isolates obtained from various clinical and foodborne samples. Immunmagnetic beads successfully captured all 74 isolates; however, only 53 isolates showed susceptibility toward the bacteriophage. Susceptible isolates were further classified into two broad groups, moderately sensitive isolates, which generated phage titer ∼107 pfu/mL (group I, n = 15), and highly susceptible isolates, which generated high phage titer ∼109 pfu/mL (group II, n = 38). We selected 15 isolates (9 from group I and 6 from group II) and individually spiked beef samples (ca. 3–8 cells/25 g beef ) to evaluate the bacteriophage-based detection system. Eight out of nine isolates from group I and all six isolates from group II were successfully detected. Pathogenic E. coli strains belonging to other serogroups (12 serogroups, 67 isolates) as well as nontarget microorganisms (n = 18) were not lysed by the bacteriophage and hence were not detected. The method is high-throughput compatible, is rapid, and can provide live culture the following day by streaking an aliquot before phage lysis on conventional selective agar media.
Introduction
F
Detection of pathogens using phages may allow the distinction of living bacteria much more efficiently than standard molecular techniques (Favrin et al., 2001). However phage-based methods may not always provide the sensitivity of a PCR approach (Goodridge et al., 1999; Favrin et al., 2003). On the other hand, measurement of ATP using a bioluminescent assay offers sensitive and rapid method in which theoretical detection limits have been predicted in the range of 103 bacterial cells (Stanley, 1989). However, when tested in complex biological matrices such as enrichments of vegetable and meat extracts, 104–105 cells were required (Thouand et al., 2008). In living cells, the enzyme adenylate kinase (AK, E. C.2.7.4.3) catalyzes the equilibrium reaction of generating two molecules of ADP from ATP and AMP in the presence of Mg++ ions. Squirrell and Murphy (1997) increased the detection sensitivity of the bioluminescent assay by focusing on AK rather than ATP as a cell marker by adding exogenous ADP to the assay mix. The released intracellular AK amplified ATP by converting exogenous ADP to ATP and AMP. The resulting bioluminescent signal was proportional to AK activity and, ultimately, to the number of cells in the sample. The lack of specificity, a major drawback in the application of AK-based bioluminescent assay, was resolved by using specific bacteriophages to lyse target cells instead of lysis buffer (Blasco et al., 1998; Wu et al., 2001). These studies were performed with laboratory pure cultures of Salmonella and E. coli O157:H7, and efficacy of AK-based bioluminescence in phage lysed cells using food matrices has not been thoroughly examined (Sun et al., 2002). In this study we used O157 immunomagnetic beads to concentrate E. coli O157:H7 cells after a short 8 h selective enrichment and utilized bacteriophage CSLO157, which is specific to E. coli O157 to lysethe pathogen. The released AK activity was measured using the luciferin–luciferase assay (Blasco et al., 1998). We reasoned that the combination of two principles, specificity (provided by immunomagnetic beads and bacteriophages) and magnification of the signal (e.g., luciferin–luciferase reaction), would enable detection of E. coli O157:H7comparable to DNA-based methods. We also took upon the task to detect E. coli O157:H7 at low contamination levels (at ∼3–8 cells/25 g) on ground beef, which was subsequently stored at 4°C for 2 days to induce cold stress. Data from the experiments involving 74 E. coli O157:H7 isolates and several other nontarget organisms (toxigenic and commensal isolates) are presented.
Materials and Methods
Bacterial strains and culture conditions
A list of E. coli O157:H7 isolates and other nontarget strains used in this study are listed in Tables 1 –4. Reference stocks of E. coli O157:H7 and other nontarget strains were stored at − 70°C in 15% (vol/vol) glycerol in LB broth. A stock culture was kept at 4°C on tryptic soy agar (Oxoid) plates and was refreshed every week. Working cultures were prepared by suspending a single colony in 10 mL tryptone soy broth (TSB; Oxoid) and then were incubated overnight (∼18 h) with shaking (200 rpm) at 37°C. Alaska diagnostics bacteriophage specific to E. coli O157 (CSLO157, obtained from The Food and Environment Research Agency, Sand Hutton, York, United Kingdom) was used.
Phage susceptibility grouping was based on plaque forming efficiency using bacteriophage CSLO157 (see Fig. 1 for details).
National Food Safety and Toxicology Center (Michigan State University).
National Food Safety and Toxicology Center (Michigan State University).
Experimental contamination of ground beef samples
Ground beef (20% fat) was purchased from a local grocery and was divided aseptically into 25-g portions in pulsifier bags (Microbiology International). Beef samples were artificially contaminated with ∼3–8 E. coli O157:H7 cells by adding 200 μL of appropriate dilutions. Viable cell counts were confirmed by retrospective spread-plating 10-fold serial dilution onto tryptic soy agar plates. Artificially contaminated beef samples were stored at 4°C for 48 h before subjecting to enrichment. All beef samples were tested for intrinsic E. coli O157:H7 by the conventional, as well as bacteriophage-based method, and were found to be negative. Each E. coli O157:H7 isolate was tested in 5 to 9 independent tests and each test included three replicate samples.
Selective enrichment protocol
To each 25-g ground beef sample, 225 mL of brain heart infusion broth (BHI; Oxoid) was added. The broth was amended with the following selective agents (μg · mL−1 final concentration): novobiocin, 11.0; cefsulodin, 11.0; vancomycin, 8.8; maltol, 4.4; and acriflavine, 10.0. The samples were pummeled in a pulsifier (Microbiology International) for 2 min and were incubated at 41.5°C for 8 h without shaking.
E. coli O157 Dynabeads capture
The protocol for the E. coli O157 Dynabeads capture (Dynal) was performed as described by the manufacturer except that 11 mL of the enrichment sample was used after it was filtered through a Alaska food filter (Alaska Diagnostics) and 20 μL of antibody-coated magnetic beads was incubated at room temperature in a Magnetic Sample Rotator (Alaska Diagnostics) for 10 min. A magnetic field was applied for 5 min, and the supernatant was carefully aspirated and discarded. Samples were washed two times with phosphate-buffered saline containing 0.1% Tween-80 (PBST). The supernatant was discarded, and the particle–bacteria complex was resuspended in 500 μL of PBST. Twenty-microliter beads were streaked on sorbitol MacConkey agar supplemented with cefixime tellurite selective supplement (CT-SMAC; Oxoid), and the remainder of the magnetic beads were subjected to E. coli O157:H7–specific phage treatment as described below.
Phage treatment and luciferin–luciferase (FastrAK™) assay to detect E. coli O157:H7
The phage treatment and luciferin–luciferase assay was performed essentially as described earlier for the FastrAK™ assay for Salmonella strains except that E. coli O157–specific phage CLSO157 was used (Patel et al., 2009; Hammack and Chen, 2010). Briefly, the suspended Dynabeads magnetic particles (from the previous step, see above) were divided into two 240-μL aliquots and placed into 2 wells of a 96-well microtiter filter plate (with 3.0 μm glass fiber/0.2 μm, Bio-Inert membrane; Europa House). The magnetic particle–bacteria complexes were washed three times with 200 μL PBST by placing the filter plate on a 96-well vacuum manifold. For each sample, one aliquot was treated with bacteriophage CSLO157 diluted to deliver 5 × 108 pfu in 100 μL, while the other aliquots received 100 μL phage diluent buffer (ATSB solution, which is PBS with 5 mM glucose and 10 mM MgSO4). The bacteriophage–magnetic bead–bacteria complex and control aliquots were incubated at 37°C for 2 h. Lysed bacteria or ATSB buffer eluate from individual wells were collected in a new plate using 96-well vacuum manifold. The plate was immediately placed in a FastrAK plate luminometer equipped with two injector-coupled tubes, one containing 10 mM ADP solution and the other with a proprietary luciferin–luciferase mixture. For each measurement, the following controls were included: bacteriophage suspension solution, adenylene kinase, unlysed magnetic bead–bacteria complex, and enrichment medium. Emitted light from wells was measured as relative light units. The relative light units from control wells (T1) were subtracted from sample wells that received bacteriophages to get the background subtracted light unit readings (T2). Each sample was assayed in triplicate and each treatment was repeated for at least three times. The data were plotted as signal-to-noise ratio (T2/T1) of relative light units.
For all statistical analyses, SigmaStat 3.0 software was used. Data were analyzed by one-way analysis of variance test or Student–Newman–Keuls method to determine statistical differences between means of treatments.
Results and Discussion
The aim of this study was to evaluate specificity and sensitivity of a bacteriophage-based assay for detection of E. coli O157:H7 in ground beef. We also optimized this assay while keeping the enrichment time at 8 h, artificial contamination level to ∼3–8 cells per 25 g ground beef, and stressing the cells at 4°C for 2 days before enrichment. Initially, we evaluated several enrichment media in addition to BHI such as buffered peptone water and modified buffered peptone water, basic listeria enrichment broth (Heller et al., 2003; Kim and Bhunia, 2008; Jasson et al., 2009), and TSB with yeast extract containing varying amounts of selective agents (novobiocin, cefsulodin, vancomycin, maltol, and acriflavine) (Jasson et al., 2009). These selective media successfully enriched E. coli O157:H7 isolates 52, 106, and 105 to a detectable threshold after 18 h incubation (data not shown). However, with a shorter enrichment period (e.g., 8 h), these media were successful only if beef samples were enriched immediately after spiking (e.g., without subjecting to cold stress). Additionally, enrichment in basic listeria enrichment broth and TSB with yeast extract media resulted in high background readings of the enzyme AK (data not shown). Among all different enrichment media examined, only BHI broth containing novobiocin, cefsulodin, vancomycin, maltol, and acirflavine generated a detectable number of E. coli O157:H7 and low background levels of adenylate kinase in the control untreated wells. The observations obtained with three E. coli O157:H7 isolates were further extended to analyze an additional 71 E. coli O157:H7 isolates (Tables 1 and 2).
Phage sensitivity of E. coli O157:H7 isolates
The ability to lyse E. coli O157:H7 is the key for this detection method; therefore, we determined phage susceptibility of all 74 isolates. Phage titer of CSLO157 initially determined on ATCC 43895 reference strain (9.5 ± 4.4 × 109) was compared with all of the other isolates to determine the sensitivity to phage-induced lysis (Fig. 1). Out of 74 E. coli O157:H7 isolates, 21 isolates were not lysed by CSO157 and produced no plaques. The remaining 53 isolates were grouped as moderately susceptible (group I, n = 15) and highly susceptible (group II, n = 38). The mean phage titer (pfu · mL−1) of group I isolates was 5.5 ± 8.5 × 107 compared with 4.2 ± 1.7 × 109 (p ≤ 0.05). All phage-resistant as well as phage-susceptible isolates (Tables 1 and 2) were determined to be of O157:H7 serogroup by latex agglutination test (Oxoid) as well as by their binding to Dynal O157 immunomagnetic beads by estimating percentage of cells recovered on CT-SMAC plates (data not shown).
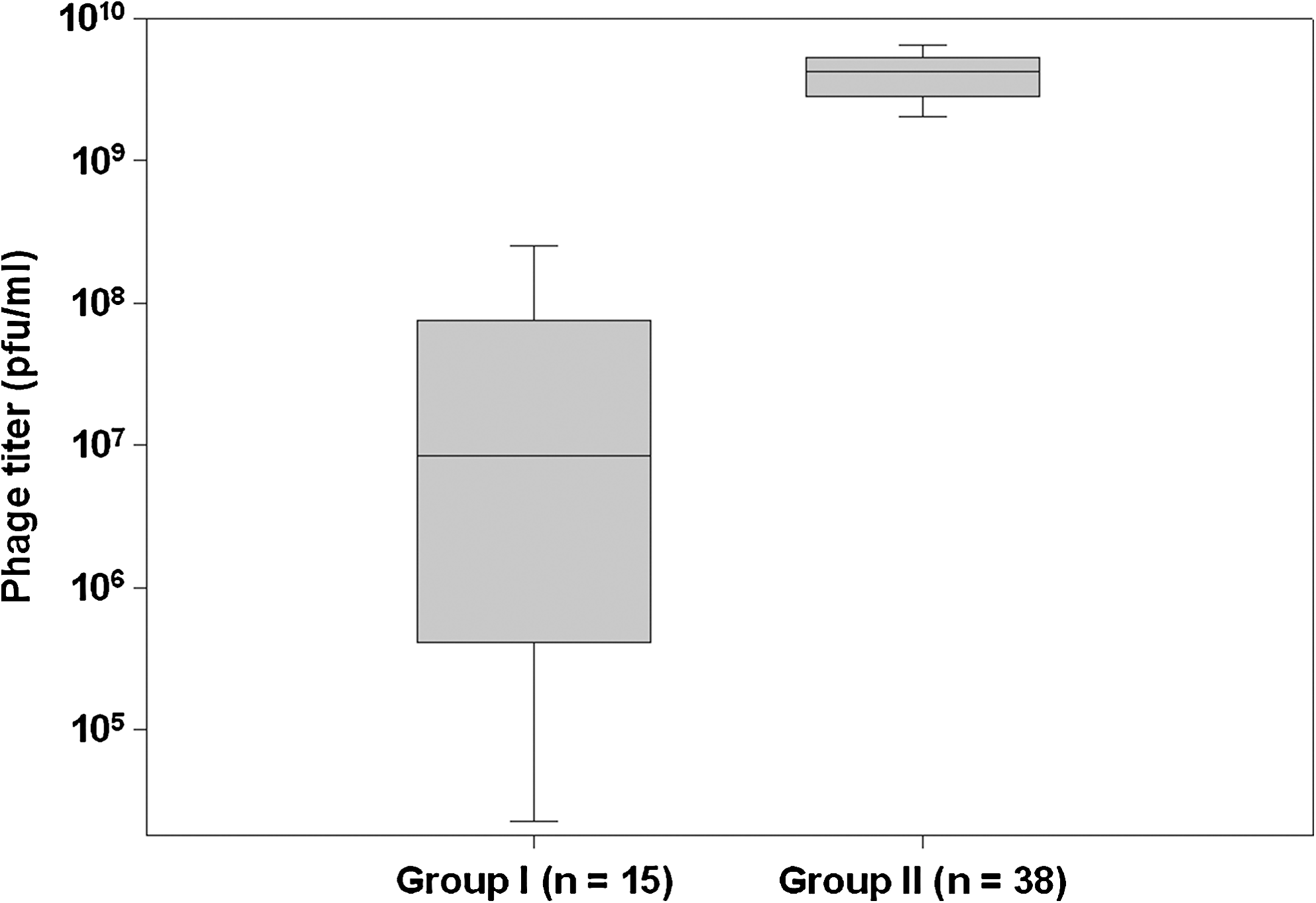
Box plot of grouping of Escherichia coli O157:H7 isolates based on phage susceptibility. Phage titer (pfu · mL−1) was determined by 10-fold serial dilution of the stock phage CSLO157. Out of 74 E. coli O157:H7 isolates, 21 isolates did not form plaques. Remaining 53 isolates were grouped based on their median plaque forming efficiency (group I, 0.85 ± 8.5 × 107 and group II, 4.2 ± 1.7 × 109; Kruskal–Wallis one-way analysis of variance on Ranks, p < 0.05). The boundary of the box closest to zero indicates the 25th percentile, a line within the box marks the median, and the boundary of the box farthest from zero indicates the 75th percentile. Whiskers (error bars) above and below the box indicate the 95th and 5th percentiles.
E. coli O157:H7–specific bacteriophages have been isolated and evaluated to control the target pathogen population in sheep and cattle feedlots (Raya et al., 2006; Callaway et al., 2008) and under vitro conditions (Niu et al., 2009). In the current study a single phage, CSLO157, specific to O157 was used. To improve the efficacy and robustness of this assay, a cocktail of phages containing diverse but O157:H7-specific phages are needed, which could be the subject for future investigations.
We tested Shiga-toxin-producing non–O157 E. coli strains isolated from clinical and foodborne outbreak studies (Bhagwat et al., 2005) for their susceptibility toward bacteriophage CSLO157. None of the serotype strains tested such as O26 (n = 13), O55 (n = 9), O111 (n = 15), or O128 (n = 5) generated any titer or visible plaques. Other serotypes tested and found to be resistant to bacteriophage CSLO157 were O44, O103, O104, O113, O119, O121, O125, O127, O133, O142, O145, and O156 (Table 3). Altogether, 67 toxigenic E. coli isolates belonging to 12 serotypes were tested and found to be resistant to the bacteriophage CSLO157. We also examined other enteropathogens such as Shigella sonnei, Shigella dysenteriae, Shigella flexneri, and Salmonella enteric serovar Typhimurium strains. Other nonpathogenic organisms tested for the phage susceptibility were Klebsiella pneumonia, Citrobacater freundi, and other commensal bacteria (n = 22) (Table 4).
Detection of group I and II isolates in artificially inoculated beef
Next we selected several group I and II isolates (Table 1) and individually spiked them on beef samples. The spiked beef samples were stored at 4°C for 2 days before enrichment. Each E. coli O157:H7 isolate was spiked in 5 to 9 independent tests and each test included three replicate samples. After 8 h enrichment at 41.5°C without shaking, samples were processed for phage lysis as outlined in the Materials and Methods section. For group I isolates, the ratios of relative light units released upon phage lysis to unlysed/untreated cells were >1.8 except for isolate A66 (Fig. 2A). Signal-to-noise ratio for isolate A66 was 0.92 ± 0.15 and was statistically similar to ratio from blank or control wells, which was 0.82 ± 0.82 (p < 0.05). Rest of the isolates showed higher signal-to-noise ratios with significant statistical difference with reference to blank or isolate A66. Detection efficacy was much better for group II isolates (Fig. 2B), where five out of six isolates had signal-to-noise ratio >10 (isolate A106 had median signal-to-noise ratio of 4.4 ± 1.4). From all samples, a portion of magnetic beads (20 μL out of 500 μL suspension after capture and wash step) were spread on CT-SMAC plates and after overnight incubation yielded typical O157:H7 type colonies.

Box plot of detection efficiency (signal-to-noise ratio, T2/T1) of E. coli O157:H7 isolates from group I (
Although isolate A66 was not successfully detected by the phage lysis method, it was probably not due to insufficient number of cells after 8 h enrichment. In all 7 tests using isolate A66, we were able to isolate O157:H7 colonies on CT-SMAC plates (data not shown). It may be noted that isolate A66 was a poor host for the phage CSLO157 and generated lowest phage titer of 1.9 ± 0.89 × 103 pfu · mL−1 among all group I strains. It is interesting to note that although isolates A99 and A114 generated titers of 3.6 ± 1.7 × 104 and 5.1 ± 2.2 × 104 pfu · mL−1 (respectively), their signal-to-noise ratio was significantly above the background (Fig. 2A). Thus, it appears that there was no effective lysis of A66 cells upon treatment with phage CSLO157 to release enough AK.
Phage susceptibility and signal-to-noise ratio
In general, group II isolates had much better signal-to-noise ratio (median ratio 14.58 ± 6.2) compared to group I isolates (5.17 ± 3.6) (p < 0.05) (Fig. 3). If we assume that higher phage titer is equivalent to efficient lysis of host cells, better detection efficacy in terms of higher signal-to-noise ratio is expected for group II isolates. There was no difference in binding efficacy of Dynal O157 immunomagnetic beads to group I and II isolates (data not shown). Secondly, both groups of isolates did generate typical E. coli O157:H7 serotype colonies on CT-SMAC plates from 20 μL aliquot of the immunomagnetic beads, and no correlation was observed in number of colonies recovered versus signal-to-noise ratio after phage lysis. This observation indicates that all isolates had reached threshold enrichment to be captured on immunomagnetic beads, and degree to which they were susceptible to bacteriophage CSO157 was reflected in the signal strength they generated upon lysis (Fig. 3).
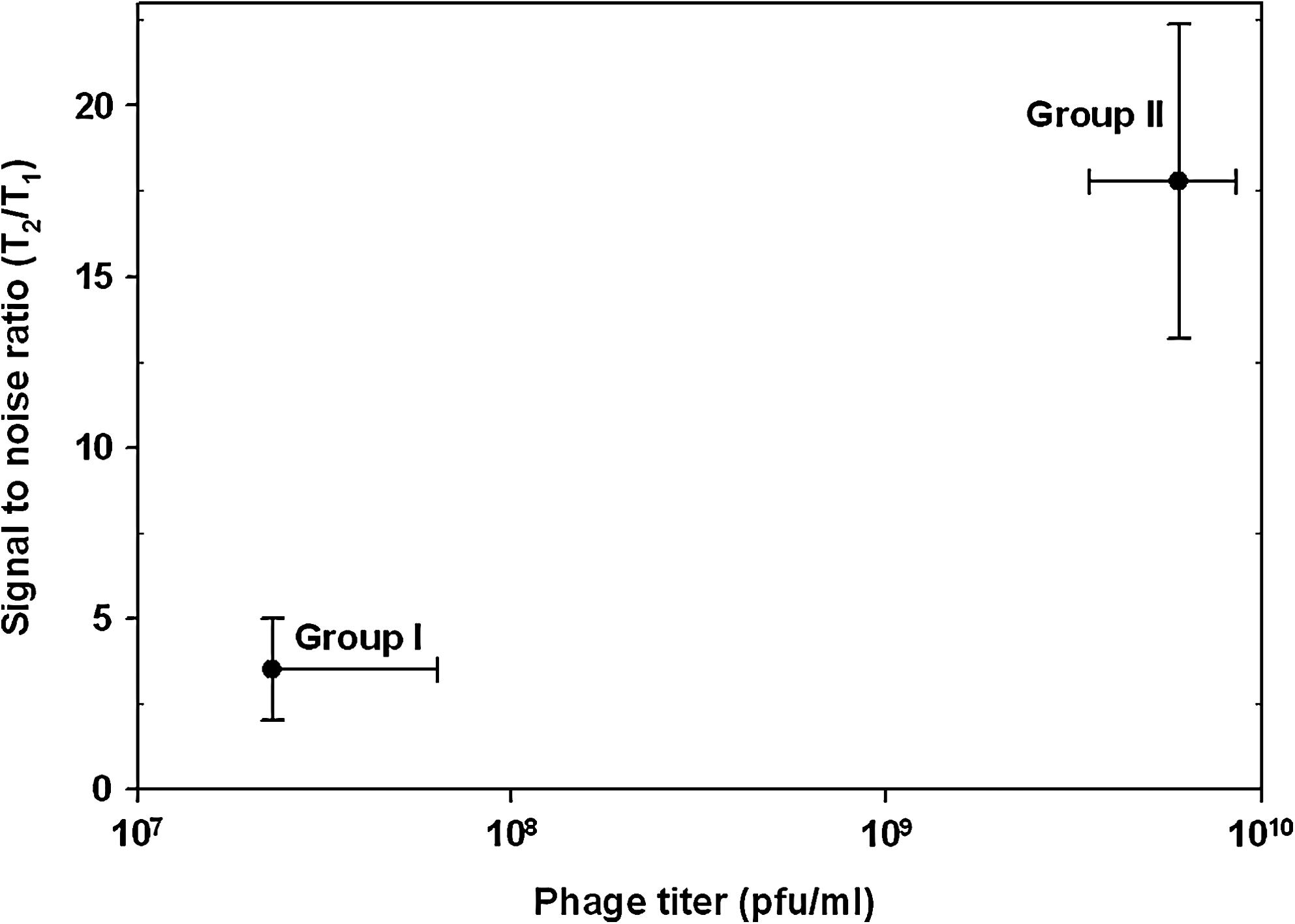
Relationship of phage titer and signal-to-noise ratio for detection of E. coli O157:H7 isolates using FastrAK phage assay. For the two-way plot, ratios from individual isolates from two groups were plotted against their phage titer values.
Conclusions
The potential of a new AK-phage assay for the detection of low numbers of E. coli O157:H7 strains in ground beef after 8 h of enrichment has been clearly demonstrated in this study. The assay successfully detected 14 out of 15 O157:H7 isolates. However, further work is needed to generate a cocktail of phages that will specifically lyse diverse range of O157:H7 isolates, as 21 out of 74 isolates were found to be resistant to phage CSLO157. A more robust assay outcome is expected using phage cocktails as opposed to single phage utilized in this study.
Disclosure Statement
No competing financial interests exist.