
Abstract
Irish breeder and intensive broiler flocks together with the corresponding poultry farm environment were sampled for the presence of Campylobacter with the aim of identifying potential sources and transmission routes of poultry flock contamination. The genetic diversity of a subset of Campylobacter isolates was examined by analysis of the flaA-short variable region (SVR). Additional discrimination for a further subset of these isolates was achieved using multilocus sequence typing (MLST) analysis. Twenty-four flaA-SVR alleles and 15 FlaA peptides were detected among 92 Campylobacter jejuni and Campylobacter coli isolates. MLST data have been determined for 30 of 92 (32.6%) flaA-SVR–typed isolates with 13 sequence types (STs) present, which were assigned to seven clonal complexes. ST45 was the most common ST identified. Vertical transmission was not found to have played a role in transmission of the pathogen to the poultry flocks. Subtyping by flaA-SVR and MLST identified the practice of partial thinning of flocks as a potential source and route of flock contamination on one broiler farm and implicated a probable source of flock contamination on another. Although there have been several studies reported in the scientific literature, the findings from this study confirmed previous studies and suggested some new transmission pathways including via transport crates. Cross-contamination from adjacent cattle is a new development, and molecular evidence of the role of transport crates in introducing Campylobacter spp. into the broiler house is a recent finding. Further, this study reports the discovery of five new flaA-SVR allele types and eight new STs. These were widespread and persistent in the poultry environment. This new knowledge may explain why despite the on-farm Campylobacter data published to date, there are still no completely effective on-farm control measures to prevent Campylobacter contamination of broiler flocks.
Introduction
Materials and Methods
Study farms and sample collection
During October 2006 to June 2007, three poultry farms, two in the Republic of Ireland and one in northern Ireland were visited. Each farm comprised two poultry houses and the flock sizes were in the region of 5000–7000 birds. Farm 1 and its corresponding breeder farms were sampled during the period October to December 2006. Farm 2 and corresponding breeder farm were sampled during the period February to March 2007, and finally, farm 3 and corresponding breeder farm were sampled during the period April to June 2007.
Sampling of cleaned poultry houses and flocks
Extensive sampling was carried out in the cleaned and disinfected broiler house prior to chick placement. In each broiler house, interior air was sampled in conjunction with the physical structure of the house and various other physical samples were taken (including feed, water, and sawdust). For all farms, samples were taken on day 1 of the following elements: interior air, feed, walls, floor, water dispenser, feeder, sawdust, internal support, windows, personnel boots. On subsequent sampling days (except day 1), the interior air was sampled on all three farms in addition to the feed (except farm 1 on the final sampling day). Flock sampling was carried out at each farm over the entire life cycle of the broiler flock (over a 5- or 6-week period). Papers lining the chick transport crates were collected from each flock on day 1, when chicks were initially introduced into the broiler house. Subsequently, 60 fresh fecal droppings (5 pooled fecal samples each containing 12 fresh fecal droppings) were collected from each house on farm 1 (on days 16, 30, and 37), farm 2 (on days 14, 25, and 37), and farm 3 (on days 14, 28, and 42). Given time constraints, fewer fecal droppings (24 fresh fecal droppings) were simultaneously collected from the second broiler house present on each poultry farm (arbitrarily designated adjacent house in Table 1) on a given sampling day.
Campylobacteraceae isolated from samples processed by direct plating, enrichment, and/or the CampyCheck method for emerging Campylobacteraceae, as appropriate to the sample type.
Twelve feces samples were amassed together in five pooled fecal samples.
Not determined or not applicable.
Environmental sampling on poultry farms
Samples from various sources (cattle feces, puddles, broiler house apron (concrete area around the house), soil, air, water supply, wild bird feces and insects) were procured prior to chick placement into the house and during the rearing period in the immediate external surrounding of the poultry house (farm) environment (Table 1). Swabs from transport equipment were also procured on one farm before the partial depopulation of the flock (farm 3 on day 34).
Isolation of Campylobacter from fecal and environmental samples
A total of 1540 samples (including 1116 fresh fecal dropping samples, 278 environment samples, and 146 crates swabs) (Table 1) were collected, transported under chilled conditions, and immediately processed in the laboratory using conventional methods and a method for the identification of emerging species of Campylobacter as previously described by Patriarchi et al. (2009). In brief, methodology based on the Horizontal Method for Detection and Enumeration of Campylobacter spp. (ISO 10272-1:2006, International Organization for Standardization, Switzerland) was used to isolate Campylobacter spp. from feces by direct plating. The CampyCheck method was used to detect emerging Campylobacteraceae as described by Lynch et al. (2007). An enrichment culture method for the detection and semiquantitative enumeration of thermotolerant Campylobacter spp. (ISO 17995:2005; International Organization for Standardization) was used to isolate Campylobacter from water samples. Air samples were subjected to direct plating and enrichment culture recovery methods as described above. A sample air MK2 (AES Laboratoire) double agar plate sampler was used to collect aerosol samples directly onto Campylobacter blood-free selective agar base (CCDA, CM0739; Oxoid) supplemented with either CCDA selective supplement (SR0115; Oxoid) or tryptone soya agar (Oxoid). Structural samples were collected from inside poultry houses, walls, structural columns, and feeders using sterile swabs premoistened with 10 mL maximum recovery diluent (MRD, CM733; Oxoid). An area of 0.1 m2 was swabbed and swabs were recovered into sterile filter bags containing 100 mL of Campylobacter enrichment broth (Lab-M) with 5% (v/v) lysed horse blood and supplement (cefoperazone, vancomycin, trimethoprim, cycloheximide [CVTC] selective supplement; Lab-M) and then samples were processed using the enrichment culture method.
Identification and speciation of Campylobacter isolates
Presumptive Campylobacter isolates were confirmed using standard biochemical procedures including the KOH, catalase (3% H2O2), and oxidase (tetramethyl-p-phenylenediamine; Sigma) reactions.
flaA-short variable region sequencing and multilocus sequence typing
Genotype analysis was carried out using flaA-short variable region (SVR) sub-typing on a subset of isolates collected from three study farms to verify the hypothesis of cross-contamination from the environment adjacent to cattle and transport crates and the broilers into the poultry houses. Selection of this subset of isolates (92 isolates) was therefore performed according to the following rationale. Approximately equal numbers of samples were selected to represent each sample type from each farm. The isolates were grouped according to farm and source, and from within these groups, samples were chosen with the objective of ensuring that every source was represented. The isolates chosen included 73 of poultry origin (poultry fecal samples), 4 of bovine origin (cattle fecal samples), 3 of wild bird origin (wild bird fecal samples), and 12 associated with environmental samples (apron, soil, and transport equipment). The multilocus sequence typing (MLST) was used solely to cross-check the flaA-SVR conclusions. MLST was performed on a subset of 30 of these isolates (poultry fecal samples, n = 21; soil, n = 2; transport equipment, n = 4; cattle faces, n = 1; broiler house apron, n = 1; wild bird feces, n = 1). MLST uses genetic variation from multiple chromosomal locations to distinguish between bacteria. In this experiment the following seven loci were chosen: aspA, glnA, gltA, glyA, pgm, tkt, and uncA. Genomic DNA was isolated using a Blood and Tissue Kit (Qiagen). The SVR of the flaA gene was amplified using primers and cycling conditions as described by Nachamkin et al. (1996). For certain isolates, successful amplification of the desired gene product was only possible when the annealing temperature of the polymerase chain reaction (PCR) was reduced to 45°C. flaA PCR products were purified using a QIAquick PCR purification kit (Qiagen). Sequencing was carried out commercially (MWG). Forward and reverse sequences obtained were aligned and trimmed to the 321-bp region corresponding to the flaA-SVR of Campylobacter type strain NCTC11168. Sequences were used to interrogate an online database (
Data analysis
Trimmed flaA-SVR sequences (including sequence types [STs]) were imported into BioNumerics, Version 5.0 (Applied Maths). A dendrogram was constructed using the unweighted-pair group with mathematical average method, which employs a sequential clustering algorithm, in which local topological relationships are identified in order of similarity, and the phylogenetic tree is build in a stepwise manner.
Results
Prevalence and distribution of Campylobacter on farms
All breeder flocks sampled in this study tested positive for Campylobacter spp. All broiler flocks became contaminated by the end of the rearing period with Campylobacter spp. being detected after 16 days in farm 1 and after 37 and 42 days in farms 2 and 3, respectively (Table 1). A high rate of isolation of Campylobacter spp. from fecal samples was observed when the birds were in the last period of rearing. Biochemical/microbiological analysis identified a total of 544 Campylobacter isolates. Seventy-nine percent of these isolates (430/544) were confirmed as Campylobacteraceae by multiplex PCR, whereas the remaining 21% (114/544) could not be speciated. Identification to species level confirmed Campylobacter jejuni as the most prevalent species (285/430) followed by Campylobacter coli (105/430) and Campylobacter lari (5/430). In addition, other members of the Campylobacteraceae family (13/430) along with Arcobacter (22/430) were identified. C. jejuni and C. coli were found in all the three farms sampled. C. lari isolates were isolated from wild bird feces samples taken from farm 3 only (Table 2). All Arcobacter isolates found were classified by performing multiplex PCR as previously described by Neubauer and Hess (2006). Arcobacter isolates were not isolated from farm 2. Despite extensive sampling of the interior of the broiler houses on all three farms prior to chick placement, no Campylobacter spp. were found. Campylobacter spp. were not detected on air outside, water supply, insects (flies), or crate papers at any location or at any time after chicks placement into the house (Table 1).
Sources of Campylobacter in the poultry farm environment
On farms 1 and 3, Campylobacter spp. were recovered. On farm 1, C. jejuni together with Arcobacter and Campylobacteraceae were isolated from cattle feces on days 16, 30, and 37. On the same farm, the apron surface and puddle around the house were positive for C. jejuni on day 37. On farm 3, the transport equipment used to carry out the partial depopulation of the flock (thinning) was found to be highly contaminated with C. jejuni and C. coli on day 34. In addition, C. jejuni was also found contaminating soil on day 14 and wild bird feces on days 0, 28, and 42. Five C. lari isolated during this study were cultured from wild bird feces on farm 3 (Table 2). The environment surrounding farm 2 was never found to be contaminated with Campylobacter spp. (Table 1).
flaA-typing and MLST
Sequencing of the SVR of the flaA gene identified five new alleles. These novel alleles were submitted on-line to the Campylobacter Fla database (
Spatial and temporal tracking of strains on farms
In general, multiple Campylobacter flaA-SVR types were present within an individual flock. Twenty-four flaA-SVR alleles (Fig. 1) and 15 FlaA peptides were detected among 92 C. jejuni and C. coli isolates. Pearson pairwise correlation and the unweighted pair group method with arithmetic mean (unweighted-pair group with mathematical average) were used to construct a dendrogram. A threshold genetic similarity of 98% was used as a cutoff coefficient value and 11 cluster genotypes were observed (Fig. 2). The spatiotemporal distribution of Campylobacter is shown in Figure 3. The breeder flock supplying farm 1 was found to be positive for Campylobacter. Several distinct subtypes were identified using flaA-SVR analysis, including allele types 34, 1022, 32, 66, and 16. On farm 1, it was apparent that the poultry houses were contaminated with Campylobacter quite early in the growing cycle of the flock; by day 16, both poultry houses were positive for Campylobacter. The predominant flaA-SVR present is allele type 1023. On the next sampling day (day 30), this allele type is no longer detectable in the flock and has been replaced by a different subtype, allele type 2, which is widely detectable in both the flock houses and environment (soil and cattle). An additional subtype was present in the flock, allele type 325. By the last sampling day (day 37), broiler flocks were contaminated by two subtypes, allele type 2 and allele type 232. Allele type 2 was also detected on the apron and in soil, whereas allele type 60 was now the unique subtype found contaminating cattle feces. On farm 1, the presence of a single genotype in a representative isolate from poultry, soil, broiler house, apron, and cattle feces was confirmed using MLST analysis. The sequence type for all isolates was identified as ST45, belonging to clonal complex ST45. Breeder flock supplying farm 2 was found to be positive for Campylobacter. Several distinct subtypes were identified using flaA-SVR analysis, including allele type 37, 255, 8, 22, and 17. On farm 2, no Campylobacter was detected in either the poultry flocks or the environment at days 14 and 25. On the final sampling day (day 37) the flock was positive for Campylobacter and three flaA-SVR subtypes were detectable, including allele types 16, 191, and 325. However, the general farm environment remains Campylobacter negative. Breeder flocks supplying farm 3 were found to be positive for Campylobacter. One distinct subtype was identified using flaA-SVR analysis, allele type 1025. On this farm, flocks were negative for Campylobacter on sampling day 14; Campylobacter was, however, detected in the soil adjacent to the poultry house and was represented by flaA-SVR allele type 15. On the next sampling day (day 28) the flock remained Campylobacter negative; however, a particular subtype of Campylobacter (allele type 1026) was detected in a fecal sample from wild birds collected adjacent to the poultry house. Uniquely on this farm, transport equipment used for the purposes of thinning the flocks was sampled when birds were 34 days old. Several subtypes were detected on the equipment, including allele types 325, 2, 21, and 191. A representative isolate from transport equipment and a representative fecal sample from poultry were subjected to MLST analysis and it was found that both possessed an identical type, ST814 belonging to clonal complex ST661 (Fig. 2).
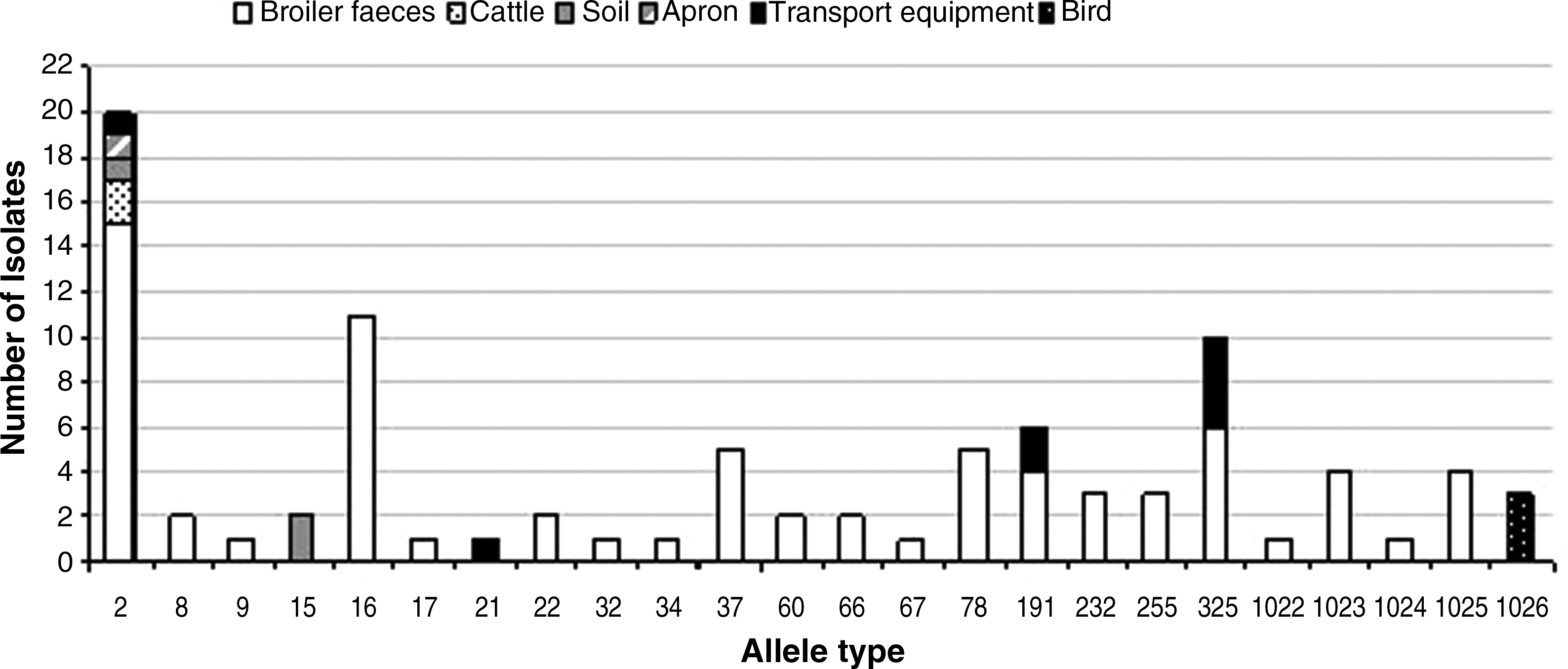
Frequency of 24 flaA-short variable region (SVR) allele types in relation to source obtained during this study. Allele type 2 was the most frequent allele type found. It was detected in broiler, cattle feces, soil, apron, and transport equipment.
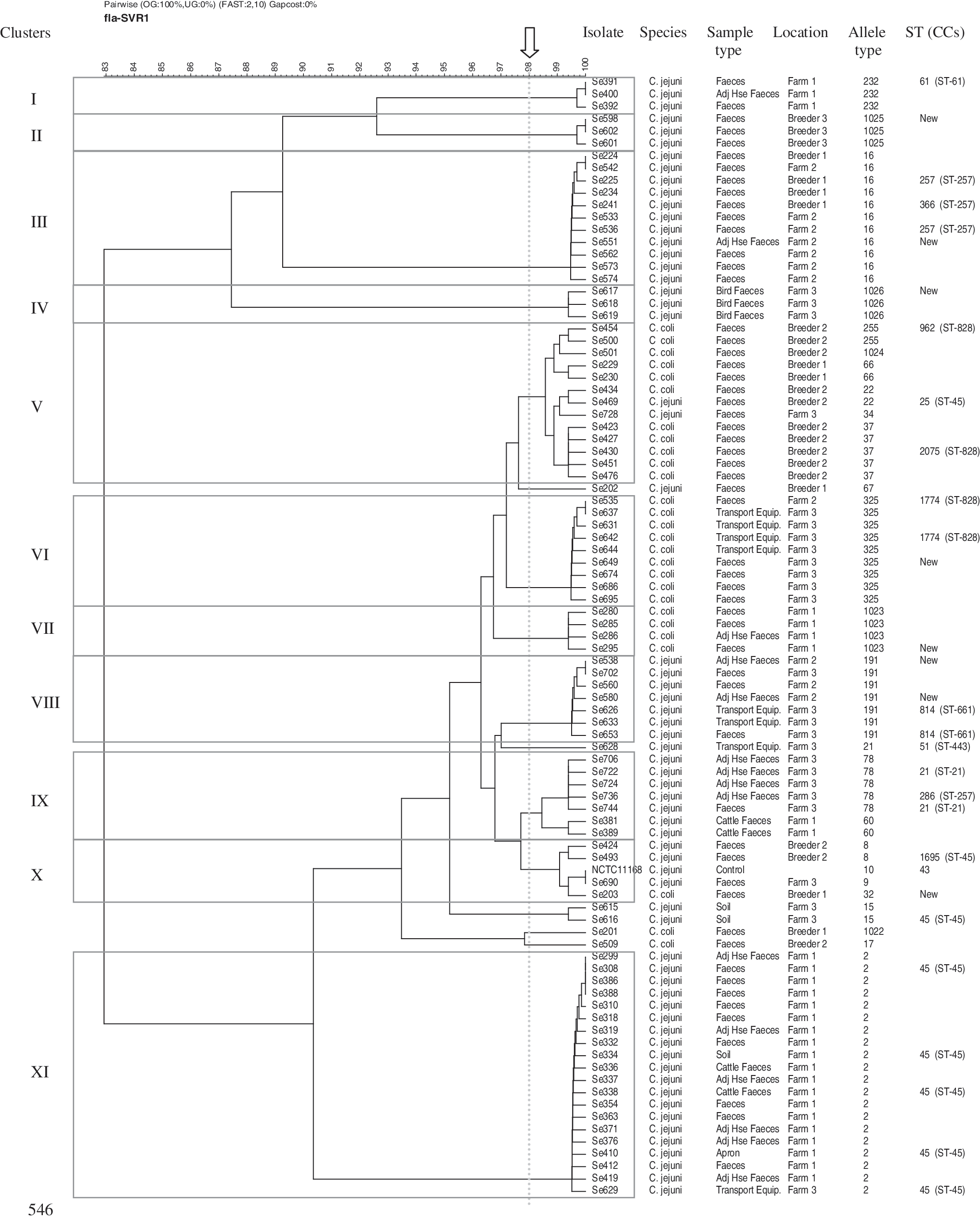
Dendrogram showing comparison of flaA-SVR sequences from Campylobacter-positive samples isolated from breeder and broiler farms and farm environments. Ninety-two Campylobacter strains were typed using flaA-SVR method. The assigned flaA-SVR allele types are indicated along with strain designations (isolate no.), the location at which the sample was taken, and the source of the sample. Strain NCTC11168 was included as a control strain in the pairwise analysis. The sequence type designation of strains that were analyzed by multilocus sequence typing is shown in the column headed “ST (CCs)” (sequence type [the clonal complex designation is shown in parentheses]) and the assigned clusters are indicated in the column on the left. The 98% similarity cutoff for flaA-SVR sequences is indicated by a dashed line.
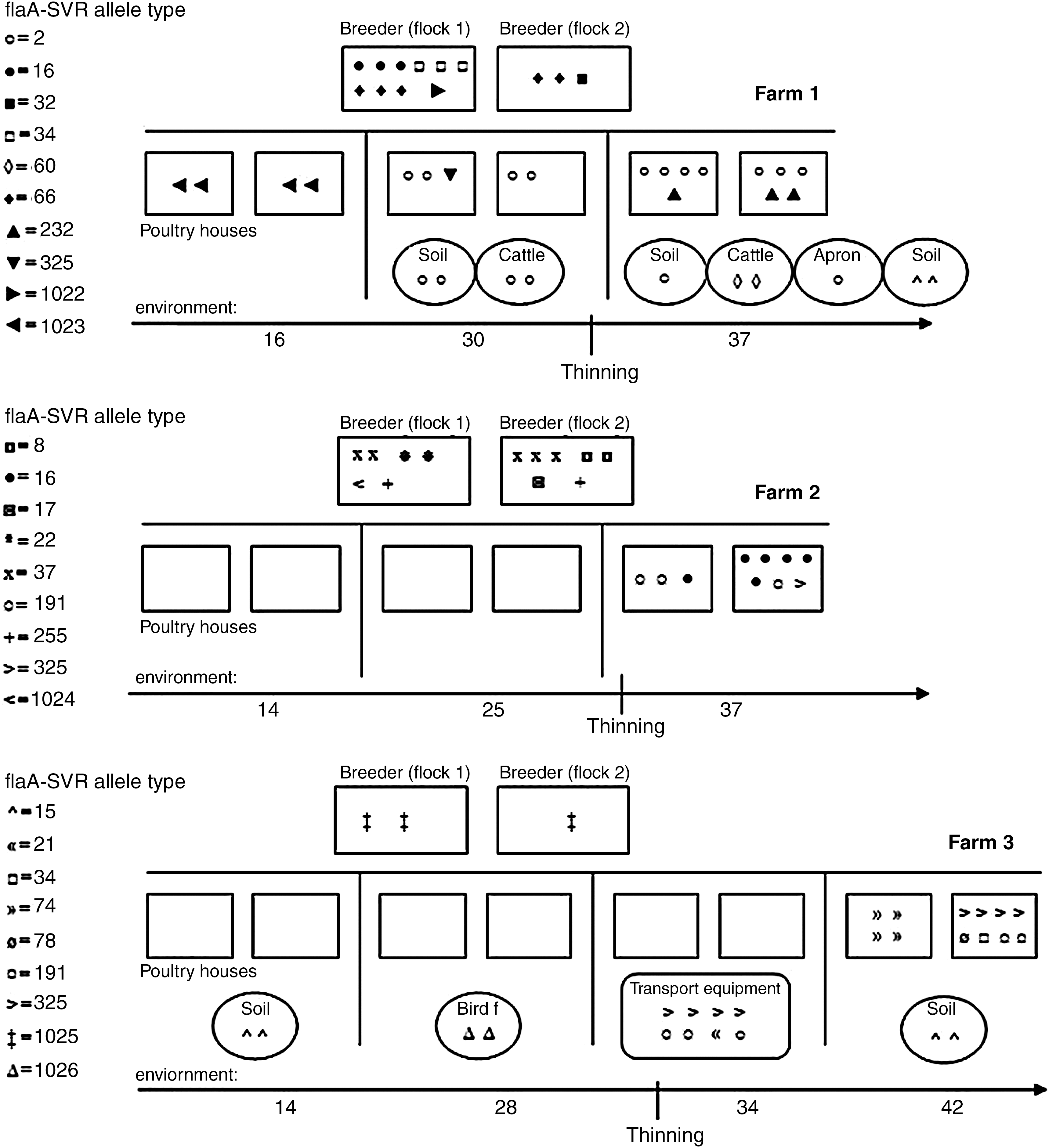
Spatio-temporal distribution of Campylobacter flaA-SVR allele types on study farms 1, 2, and 3.
Discussion
This study aimed to identify sources and transmission routes of Campylobacter in intensive housed poultry flocks from two poultry production companies in Ireland. The poultry farm environment was extensively targeted as a potential source of contamination and as such was sampled over the life span of the poultry flock on each farm. With the aim of investigating the possible routes and dynamics of transmission from identified environmental sources, that is, soil, livestock feces, and transport equipment, a subset of isolates were subjected to molecular subtyping analysis.
Molecular typing
Sequencing of the SVR of the flaA gene has been shown to be useful in determining the relationships between Campylobacter strains (Meinersmann et al., 1997). This approach is useful for short-term epidemiological studies and has been successfully applied to Campylobacter strains from a variety of sources including farm environments and slaughter and processing facilities and indeed to isolates recovered from individuals diagnosed with campylobacteriosis (Hiett et al., 2007; Callicott et al., 2008; Simmons et al., 2008). Additional discrimination power was conferred by the MLST typing method on a subset of our isolates. We have been able to exploit the combined discriminatory power of flaA-SVR subtyping and MLST to establish transmission routes of Campylobacter in selected intensive poultry flocks.
Genetic diversity
Our study found genotypic difference among Campylobacter isolates that were colonizing single flocks within each farm sampled. Multiple allele types were detected in a single flock. The dynamic of genotypic diversity among Campylobacter isolates from each farm sampled varied over time. Genetic diversity was found to increase during the months April to June, suggesting that certain genotypes occur more frequently during the spring/early summer seasons. Clonal complexes ST661, ST443, and ST21 were only detected during the months April to June on farm 3, whereas clonal complexes ST45 and ST257 were found in all the three farms sampled in the different seasons. A previous study (Sopwith et al., 2008) reported C. jejuni multilocus sequence type ST45 occurrence in surface waters as associated with early summer onset.
Vertical transmission
Campylobacter was identified in all of the breeder flocks sampled on all farms. Diverse genotypes were identified using flaA-SVR analysis; however, none of the genotypes were subsequently identified in any of the broiler flocks sampled on any of the study farms. This observation suggests that vertical transmission is not occurring between breeder and broiler flocks on the study farms. This is in line with other studies that support the hypothesis that vertical transmission does not play a significant role in the contamination of broiler flocks with Campylobacter (Callicott et al., 2006). Additionally, our observations lend further support to the hypothesis that attempts at reducing the carriage levels of Campylobacter among breeder flocks may have little impact on the potential for corresponding broiler flock contamination. Future intervention protocols should be devised and modified with this caveat in mind.
Biosecurity
The presence of a strict hygiene barrier throughout the life of the broiler flock appears to be a critical factor to avoid Campylobacter colonization. In Ireland, in line with other European producers, flocks are usually thinned between 4 and 5 weeks of age. Although not the sole source of Campylobacter, the practice of flock thinning has been previously shown to be a risk-laden process in the poultry production chain and represents a particularly recalcitrant step in biosecurity procedures. This practice has been recognized as a stressful procedure for poultry and can result in widespread shedding of Campylobacter in an intensive flock poultry house (Whyte et al., 2001). Further, environmental stress can result in modulating Campylobacter virulence having implications in the pathogenesis of campylobacteriosis (Mihaljevic et al., 2007). An ideal situation would be discontinuation of the practice of thinning, although this would have financial impacts for poultry producers. This study has provided additional evidence that thinning is a major risk factor for contamination of a previously Campylobacter-free flock. On our study farms, four flocks, two on each farm, were Campylobacter negative until the thinning process was carried out. After thinning, all flocks were Campylobacter positive. Molecular subtyping analysis successfully tracked on farm contamination routes and identified persistent strains. Farm-level data on an individual farm-by-farm basis could help fine-tune biosecurity measures already in place and, in the case of breaches, identify the most successful intervention point(s). When molecular epidemiological data from each study farm is examined it is apparent that a distinct pattern of distribution and strain transmission is occurring. The dynamics of Campylobacter over the life span of the study flocks differed, with poultry on one farm (farm 2) remaining negative for the presence of Campylobacter until the final sampling day. Conversely, flocks on farm 1 were already positive for the presence of Campylobacter by day 16 of the sampling schedule. This observation lead us to suppose different levels of compliance with biosecurity measures among the study farms. Personnel on farm 1 may not have been fully observant of biosecurity restrictions in place. On farm 1, both flocks were heavily contaminated with a particular subtype designated flaA-SVR allele type 2 by day 30 in the sampling schedule. Significantly, this subtype is also widely present in the poultry farm environment and detectable in cattle feces and soil and on the broiler house apron on the same sampling day. Confirmation of the clonal nature of the strains was obtained using MLST analysis. The strains isolated from poultry, the poultry house apron, soil, and cattle feces had identical sequence types (STs). This particular strain appeared to have a propensity for survival in the farm environment and the poultry house, suggesting strain adaptation to different environmental niches, thus presenting particular challenges for biosecurity measures in place. Comparative genome analysis could provide additional insight into this particular strains' ability to persist. Thus, elucidation of the temporal characteristics of genotypes from different sources in combination with comparative genome analysis may help clarify the role of particular genotypes in persistence and spread.
Thinning
Significantly, on farm 3, Campylobacter was isolated from transport equipment that was used in the thinning process. When molecular subtyping techniques were applied to these isolates, it was found that the strains were indistinguishable from those in the poultry flock on the subsequent sampling day (day 42). Additionally, MLST analysis of the strain designated flaA-SVR allele type 191 isolated from poultry feces and transport equipment revealed that the sequence type was ST814, confirming the clonal nature of these strains. Studies have shown that transport equipment can be contaminated with Campylobacter even following washing procedures (Slader et al., 2002). A more recent study by Allen et al. (2008) found an association between strains that occurred at specific sampling sites and those isolated from flocks postthinning and the authors speculate that strains may have spread between their study farms, which shared transport vehicles and/or personnel. Despite the fact that the three study farms were affiliated with different production and processing operations, one of the genotypic clusters (cluster XI) was commonly detected from samples originating from different origins. The findings from this study arguably reveal that current truck wash and crate module disinfection processes implemented at the poultry-processing facilities are not adequate to prevent Campylobacter contamination of flocks, at least in the case of two farms (farms 2 and 3). Additionally, this study once again highlights the importance of crate and transport equipment decontamination, steps that would have a minimal financial impact on the industry and could have important impacts on public health. Genotyping established that Campylobacter isolates with ST45 clonal complex were especially prevalent both in flocks and in the farm environment.
Cattle
Clones found within the flock were widely distributed in the surrounding farm environment and in cattle fecal samples. We suggest that in addition to the risk-laden practice of flock thinning, the presence of cattle feces represent a major and underestimated risk factor for contamination of flocks with Campylobacter. This suggestion finds accordance with other recent studies that found identical Campylobacter genotypes in cattle and poultry (Ragimbeau et al., 2008). Indistinguishable strains were identified in cattle feces, apron, soil, adjacent poultry house, and transport equipment and inside the poultry houses. Therefore, this study strongly suggests that the environment, that is, horizontal transmission, may be an important source of poultry flock contamination and emphasizes the importance of biosecurity on intensive poultry farms. A recent report describing a longitudinal study of C. jejuni in cattle herds using MLST has highlighted the significance of cattle and beef products as sources or transmission routes for Campylobacter infections (Kwan et al., 2008). Another study observed matching genotypes of Campylobacter contaminating poultry farms and other farms such as cattle, emphasizing the important role of cattle in the poultry flocks colonizing process (Zweifel et al., 2008). Although the directionality of spread from cattle to poultry cannot be definitively confirmed in this study, it is clear that Campylobacter strains found in cattle were identical to those found in poultry by flaA-SVR and MLST analysis.
It is most likely that the strains were introduced into the poultry flocks on the boots or clothing of farm personnel who may not have observed the hygiene barrier, that is, boot dipping or a change to dedicated footwear before entering the poultry house. Analysis of all three study farms suggested that on farms where there was no contamination of the flock prior to thinning (farms 2 and 3), these simple practices were probably observed. Further analysis of on-farm transmission pathways will facilitate modification of existing intervention strategies that will aggressively target specific locations on poultry farms. Surveys are continuing on a national basis and future results should provide critical information that will facilitate a reduction in the incidence of Campylobacter at broiler flock level.
Footnotes
Acknowledgment
This work was funded by safefood, the Food Safety Promotion Board (04-RESR-04).
Disclosure Statement
No competing financial interests exist.