
Abstract
Aeromonas hydrophila is the most well known of the six species of Aeromonas, which has been linked to two groups of human diseases: septicemia and gastroenteritis. Reference strain ATCC 7966 and biofilm strains TUB19, TUB20, and TUB21 were investigated for their ability to form biofilm in vitro (after 48 h on polystyrene surface) and on the surface of two green-leafy vegetables, cabbage and lettuce (after 1, 2, 4, and 24 h). Attachment strength (S R) of these strains to the vegetable surface was also measured in the same time intervals. The ATCC 7966 and TUB19 had high ability to form biofilm in vitro compared with TUB20 and TUB21 in full strength tryptone soy broth or under starvation conditions in diluted tryptone soy broth (1:20, v/v). Cell surface hydrophobicity of the biofilm strains was lower than that of the reference strain. The biofilm of all tested strains on polystyrene surfaces differed from that on the vegetable surfaces. All strains studied rapidly attached to both green leafy vegetables (after 1 h). S R and cell populations (loosely and strongly attached cells) significantly (p < 0.05) increased with contact time; however, no significant (p > 0.05) differences in cell populations were recorded after 4 and 24 h. The highest S R and cell population (log CFU cm−2) were recorded by TUB19. In conclusion, the use of A. hydrophila strains isolated from environmental biofilm samples may be more useful for understanding biofilm formation on green-leafy vegetables than the reference or laboratory strains. The attachment of A. hydrophila was significantly affected by the surfaces of green-leafy vegetables. Further studies are required to improve our understanding of the interaction between human microbial pathogens and surfaces of raw vegetables.
Introduction
Aeromonas hydrophila is a Gram-negative, facultative anaerobic, motile, rod-shaped bacterium, found in fresh, marine, chlorinated, and unchlorinated water (Asha et al., 2004; Scoaris et al., 2008).
Aeromonas may be responsible for the many gastrointestinal infections (∼40%) for which no etiological agent is found (McMahon and Wilson, 2001). A greater risk of infection is reported in young children, elderly people, and immunocompromised patients (Janda, 1991). A. hydrophila is characterized by its resistant to chlorine and refrigeration (García-Gimeno et al., 1996). The public health significance of the large numbers of Aeromonas spp. that can develop in water is unclear (Scoaris et al., 2008). Foods from which A. hydrophila was isolated were most likely contaminated by water, animal feces, or foodhandlers (Kirov, 1993). A. hydrophila had been linked with contamination of fresh vegetables, including parsley, spinach, celery, alfalfa sprouts, broccoli, cauliflower, and lettuce (Callister and Agger, 1987; Berrang et al., 1989; García-Gimeno et al., 1996). The behavior of biofilm formation by A. hydrophila in vegetable salads varies according to vegetable type (García-Gimeno et al., 1996), source of isolation (Ascencio et al., 1995; Asha et al., 2004; Khajanchi et al., 2009, 2010), hydrophobicity of bacterial cells and substratum (Danese et al., 2000; Faille et al., 2002), biological factors (Merino et al., 1999; Ukuku and Fett, 2002), and environmental conditions (Flemming, 1993).
There is a limited number of investigations interested on the adhesion and biofilm formation on the surface of fresh vegetables. The influence of strain and growth temperature of Listeria spp. on the attachment to cabbage was studied by Ells and Hansen (2006). The attachment efficiency was dependent on specific strains and was not related to species or serotypes. However, the attachment strength was increased with contact time and showed a temperature-dependent response only in the early stages of exposure. Bacterial attachment to plant cells or food contact surfaces is a crucial first step to causing infection or disease (Santiago et al., 1999). Firmly attached microorganisms are a potential contamination problem to the food because of their increased resistance to a wide range of sanitizers (Oh and Marshall, 1995). Boyer et al. (2007) stated that the ability of Escherichia coli O157:H7 to form biofilm on lettuce was affected by cell-surface hydrophobicity. Patel and Sharma (2010) mentioned that Salmonella attachment to both cabbage and lettuce surfaces increased with time. Moreover, the overall attachment strength of Salmonella was significantly lower on cabbage followed by Iceberg and Romaine lettuce.
The present study was designed to investigate the ability of three A. hydrophila strains isolated from biofilm samples obtained from drinking water distribution networks to form biofilm on green-leafy vegetables compared with the ability of a reference strain of A. hydrophila. Attachment strength of these strains to cabbage and lettuce surfaces was also studied.
Materials and Methods
Bacterial strains
In addition to the reference strain A. hydrophila ATCC 7966, three A. hydrophila strains (TUB19, TUB20, and TUB21) were used in this study. These strains were isolated from biofilm samples collected from a drinking water distribution network in Taif city, Kingdom of Saudi Arabia. These strains were previously identified by partial sequencing of the 16S rRNA gene. The sequences were deposited to the EMBL/GenBank/DDBJ databases under accession nos. FN568022, FN568023, and FN568024, respectively (Elhariry et al., submitted article). Working stock cultures were maintained in 15% v/v glycerol/tryptone soy broth (TSB) at −70°C. All strains were reactivated and subcultured on TSB at 30°C, and stored at 4°C on tryptone soy agar (TSA) slants.
Determination of hydrophobicity
Percentage of hydrophobicity was determined by the microorganism adhesion to hydrocarbon test (Rosenberg et al., 1980) with xylene as the hydrocarbon (Gary et al., 2003). An overnight grown culture (at 30°C for 18 h) was centrifuged at 4600 g for 10 min and the pellet was resuspended in phosphate-buffered saline (PBS, pH 7.4) to an A 600 nm of ∼1, to eliminate any cell concentration effects. Samples of each suspension (3 mL) were added to 3 mL xylene, mixed on a vortex mixer for 1 min, incubated at 30°C for 10 min, and then vigorously remixed again on a vortex mixer at ambient temperature for 2 min. A 600 nm of the aqueous phase was measured after standing at ambient temperature for 20 min. The percentage of hydrophobicity (%h) was determined from the absorbance of the initial bacterial cell (Ai) and the absorbance of the aqueous phase after mixing with xylene (Af) using the following equation: %h = [(Ai – Af) × 100]/Ai. For statistical analysis, three independent experiments were carried out.
In vitro biofilm formation
The commonly used microtiter plate method for determining bacterial adhesion to plastic surface was applied in the present study (Rode et al., 2007). Briefly, the wells of sterile 96-well polystyrene microtiter plates (Falcon plastics; Becton Dickinson Labware) were filled with 230 μL of TSB. About 20 μL of each cell culture was added into each well (eight wells for each strain). Plates were incubated under static conditions at 30°C for 48 h. The negative control wells contained TSB only. The contents of the microtiter plates were poured off and the wells were washed three times with 300 μL of PBS. The remaining attached bacteria were fixed with 250 μL of methanol per well. After 15 min, microtiter plates were emptied and air-dried. The microtiter plates were stained with 250 μL per well of 1% crystal violet used for Gram staining (Merck) for 5 min. The excess of stain was rinsed off by placing the microtiter plates under running tap water. After the microtiter plates were air-dried, the dye bound to the adherent cells was extracted with 250 μL of 33% (v/v) glacial acetic acid per well. The optical density (OD) of each well was measured at 570 nm using a UV/Visible spectrophotometer (6105-Jenway). To study the ability to form biofilm under starvation conditions, the same procedure was repeated using TSB diluted with sterilized distilled water (1:20, v:v). For statistical analysis, three independent experiments were carried out per medium.
Attachment to cabbage and lettuce discs
Preparation of inoculum
The cells of each strain were harvested from an 18-h overnight culture in TSB at 30°C and 100 rpm by centrifugation (5000 g for 15 min at 10°C). The harvested cells were washed twice in PBS (pH 7.0 ± 0.2) and then resuspended in the same buffer to obtain an A 600 nm of 0.1. A final density of ∼106 CFU mL−1 was achieved by diluting the suspension in PBS tempered to 10°C (Ells and Hansen, 2006). Verification of cell densities were performed by spread plating techniques on TSA.
Preparing produce surfaces
Two leafy-green vegetables, cabbage (Brassica oleracea sp. capitata) and lettuce (Lactuca sativa) heads, were obtained from a local retail store in Taif city, Kingdom of Saudi Arabia. On the same day of purchase, produce discs were prepared for the attachment assay according to the method described by Patel and Sharma (2010). For each head of produce, the two outermost leaf layers were removed aseptically and discarded. Disc-shaped pieces were cut using a sterilized 2-cm-diameter cork-borer and stored in empty 100 × 15 mm Petri dishes (Fisher Scientific) with distilled water to preserve humidity. Lettuce or cabbage pieces were stored at 4°C until the time of inoculation and used within 48 h of their preparation. The initial background flora (CFU per cm2) was determined by pour plate count technique onto TSA after incubation for 48 h at 30°C.
Inoculation of cabbage and lettuce discs
The method described by Patel and Sharma (2010) was used for inoculation with some modifications. The prepared bacterial suspension (11 mL) of each strain was transferred into a sterile Petri dish. Discs of lettuce or cabbage were aseptically placed in A. hydrophila suspensions and incubated at 10°C for 24 h without agitation. The lower assay temperature was chosen to reduce the effect of bacterial growth (Garrood et al., 2004). At specific time intervals (1, 2, 4, and 24 h), pieces were removed from suspensions using sterile forceps and dipped in a Petri dish containing 11 mL sterile PBS for 2–3 s to remove residual cells carried over from the inoculum. Populations of loosely and strongly attached bacteria were determined as described by Ells and Hansen (2006) and modified by Patel and Sharma (2010). Discs were transferred into a sterile 50 mL centrifuge tube (Fisher Scientific) containing 25 mL sterile PBS with 0.1% Tween 20 (Fisher Scientific) and vortexed for 20 s to remove loosely attached A. hydrophila cells. To recover populations of strongly attached A. hydrophila, the vortexed pieces were transferred into a 50 mL centrifuge tube containing 25 mL buffered peptone water and sonicated for 30 s using an homogenizer (PolyTron® PT-2500 E; Kinematica). The homogenizer was sterilized in 70% ethanol between each sample and rinsed twice in sterile distilled water to remove residual ethanol. Enumeration of A. hydrophila was carried out by spread-pleating the appropriately diluted homogenates of PBS containing loosely attached cells or buffered peptone water containing strongly attached cells on starch- ampicillin agar (Palumbo et al., 1985). After incubation (28°C for 24 h), the populations of loosely and strongly attached A. hydrophila obtained at each sampling period were expressed to log CFU cm−2 for disc surfaces. A. hydrophila were confirmed by flooding the plates with iodine solution (4–5 mL), and amylase-positive colonies (those having a clear zone surrounding the colony) were scored as presumptive A. hydrophila. Pieces of discs inoculated in sterile water served as controls. For statistical analysis, three independent experiments were carried out per each produce type.
Attachment strength
The attachment strength (S R) was calculated as described by Dickson and Koohmaraie (1989). S R value represents the percentage of the total population of bacteria associated with produce surface that were strongly attached to produce surface [S R = (strongly attached bacteria)/(strongly attached bacteria + loosely attached bacteria)].
Statistical analysis
The statistical analysis procedure suggested by Patel and Sharma (2010) was applied in the present study. The data obtained from three replicates were analyzed by a two-way analysis of variance using “Proc Mixed” (SAS 8.2) for interaction effects of the strains, produce, and sampling period. In all cases, the level of statistical significance was of p < 0.05.
Results
Hydrophobicity and in vitro biofilm formation
Cell surface hydrophobicity of A. hydrophila, illustrated in Figure 1, is expressed as hydrophobicity percentage. In general, the reference strain A. hydrophila ATCC 7966 showed the highest hydrophobicity (32.2% ± 1.6%). The hydrophobicity percentage of the other biofilm strains TUB19, TUB20, and TUB21 ranged from 24.7% ± 1.4% to 27.6% ± 1.9% without significant (p > 0.05) differences between the hydrophobicity of these strains. However, there was a significant difference (p < 0.05) between the hydrophobicity of the reference strain and the studied biofilm strains. The ability of A. hydrophila to form biofilm on the polystyrene surface in full strength and diluted TSB (1:20) was examined (Fig. 2). In all studied strains biofilm formation was significantly (p < 0.05) affected by diluting the growth medium. They showed significantly less biofilm formation under starvation conditions in the diluted TSB compared with full strength TSB. Insignificant (p > 0.05) deference was recorded in biofilm formation of the reference strain ATCC 7966 compared with that of the biofilm strain TUB19, when the full strength TSB was tested. However, biofilm formation by the biofilm strains TUB20 and TUB21 was lower than that obtained by the reference strain ATCC 7966 and the biofilm strain TUB19. Similar behavior of biofilm formation was noticed by all studied strains in the diluted TSB, although OD 570nm values were lower than those obtained in the full strength TSB (Fig. 2).
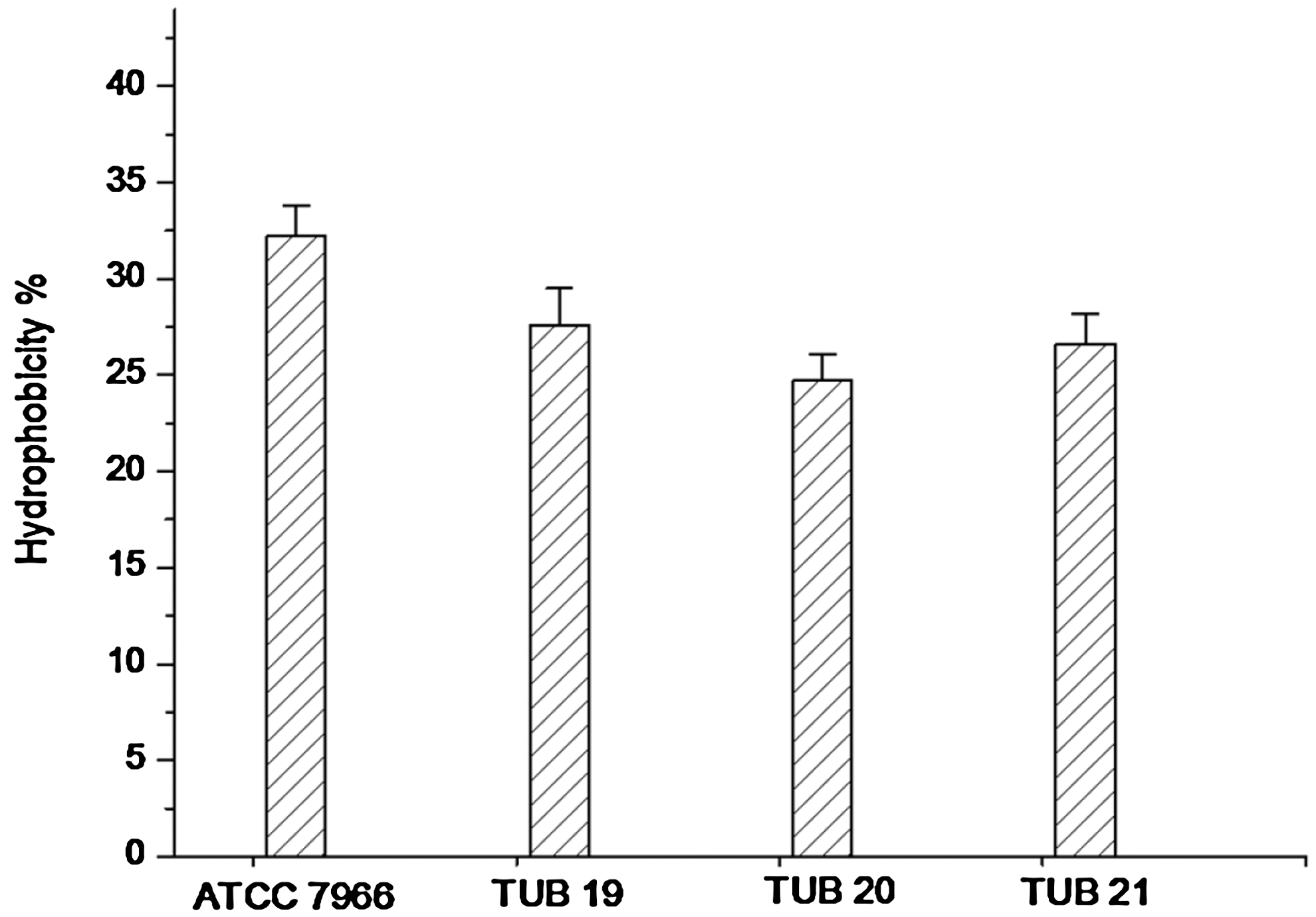
Hydrophobicity of Aeromonas hydrophila strains (the reference strain ATCC 7966 and biofilm strains TUB19, TUB20, and TUB21) determined using the microorganism adhesion to hydrocarbon test. Error bars are the standard deviation from three replications.
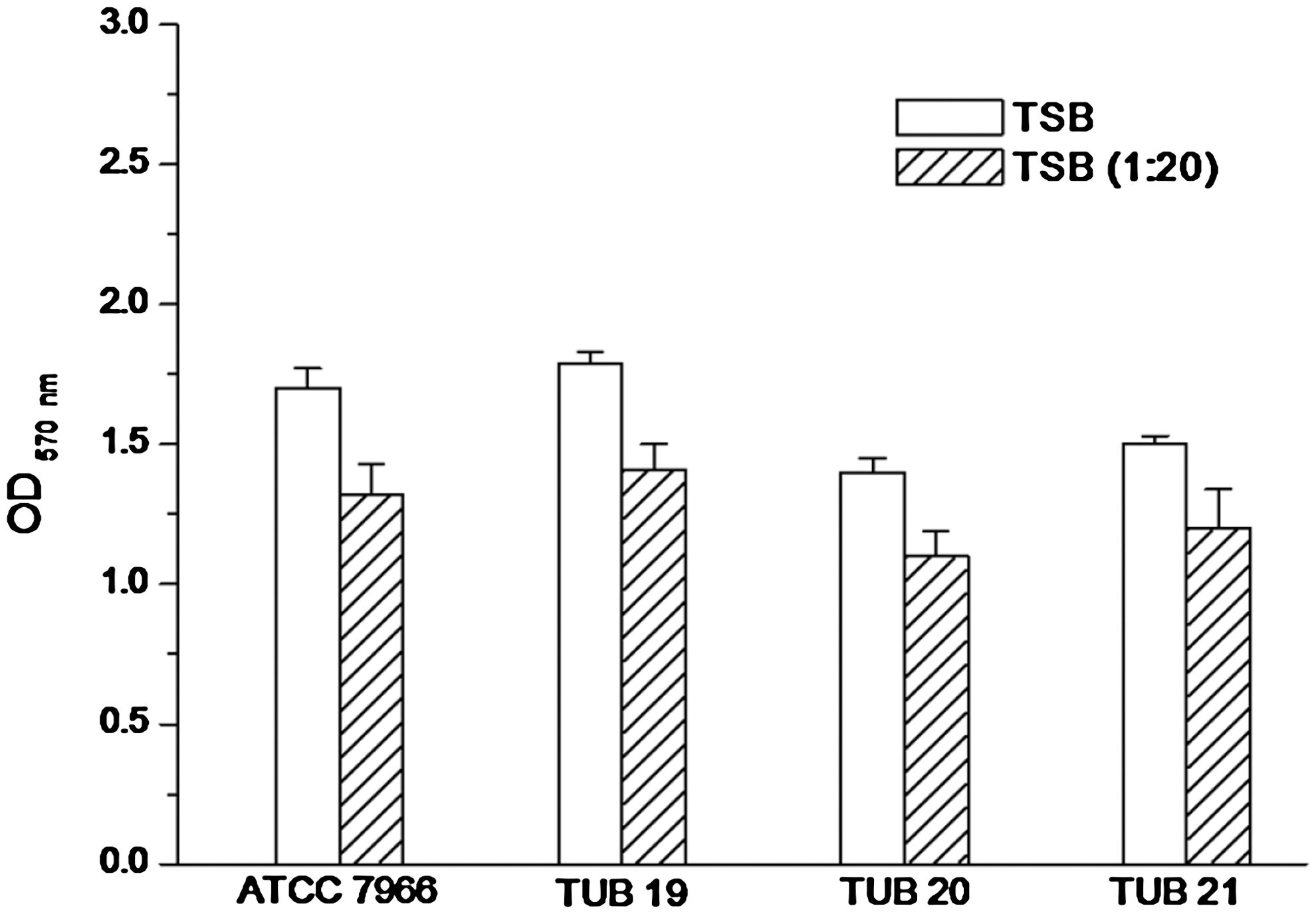
In vitro biofilm formation by A. hydrophila strains (the reference strain ATCC 7966 and biofilm strains TUB19, TUB20, and TUB21) on polystyrene. Biofilm formation was measured as optical density (OD) at 570 nm after 48h in tryptone soy broth (TBS) and diluted TBS (1:20). Error bars are the standard deviation from three replications.
Attachment of A. hydrophila cells to produce surfaces
Preliminary plating studies on TSA demonstrated that most cabbage and lettuce discs were devoid of culturable natural background flora or, if present, the total bacterial counts were very low (not > 2 CFU cm−1) and had no significant statistical effect at p = 0.05. For more adequate results, the enumeration of A. hydrophila was carried out on selective medium (starch-ampicillin agar) (Palumbo et al., 1985). The results for the loose and strong attachment of A. hydrophila cells to cabbage and lettuce discs over time are expressed as log CFU cm−2 and illustrated by Figure 3A and B. In general, A. hydrophila attachment to lettuce was significantly greater and rapid than the attachment to cabbage. Number of attached A. hydrophila cells attached to lettuce surface after 1 h was in the range of 1.6–2.4 log CFU cm−2 compared with 1.45–1.84 log CFU cm−2 for the cabbage surface. After 2 h, more cells of the studied biofilm strains were attached to the lettuce surface compared with that of the reference strain. However, no significant (p > 0.05) difference between attached cells of all studied strains was found when their biofilm formation ability on cabbage surface was studied. Although there were variations in the recorded log CFU values for each strain, no significant difference between these values was found by extending the incubation time from 4 to 24 h (Fig. 3A, B). Remarkably, the biofilm strain TUB19 had the greatest ability to form biofilm compared with the other studied strains (reference or biofilm strains). After 24 h, the attached biofilm cells ranged from 2.85 to 3.95 log CFU cm−2 and 2.73 to 3.75 log CFU cm−2 on lettuce and cabbage surfaces, respectively, with no statistical significant differences between attachments to the two surfaces. However, these values were significantly (p < 0.05) higher than that recorded by the reference strain.
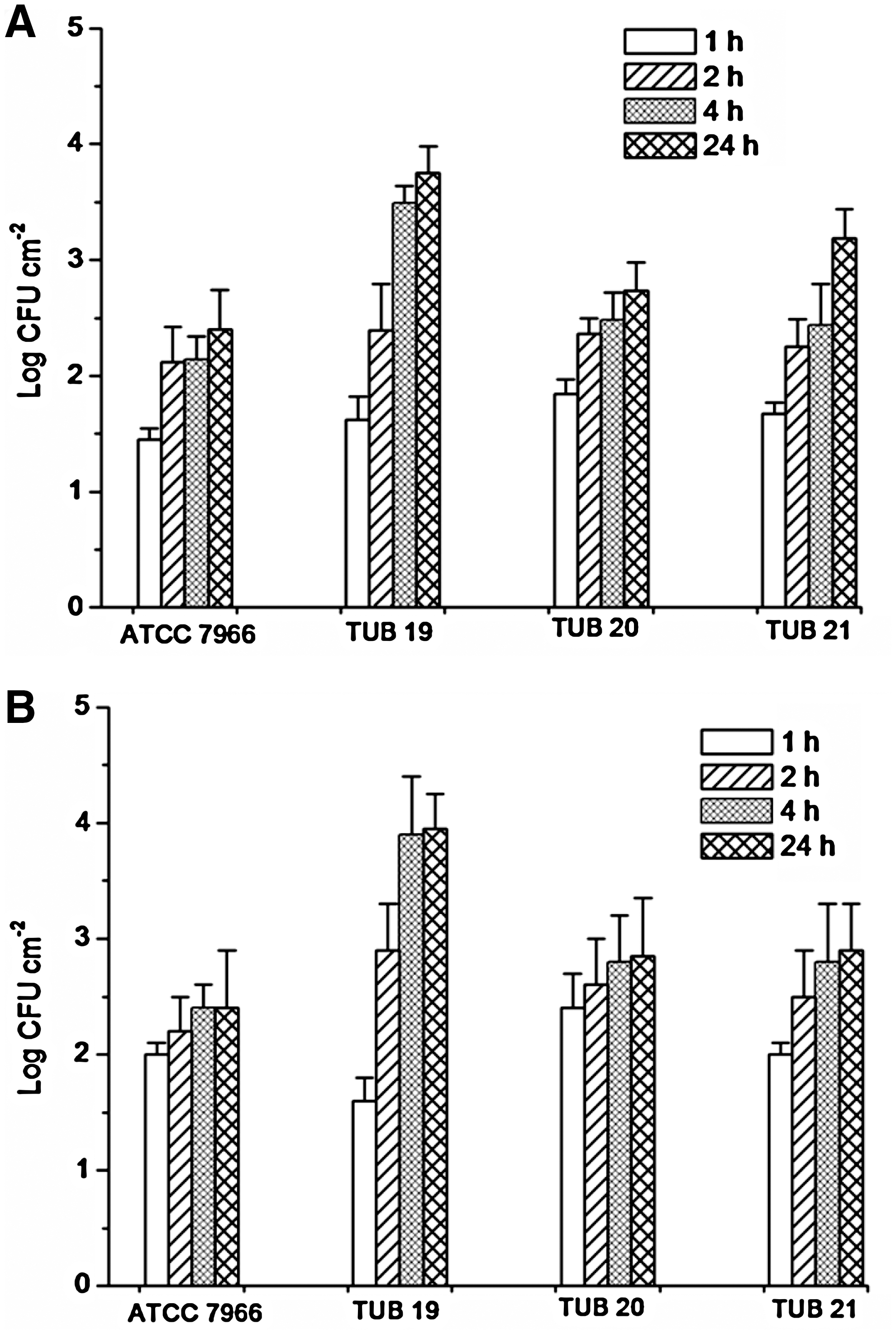
Attachment of A. hydrophila strains (the reference strain ATCC 7966 and biofilm strains TUB19, TUB20, and TUB21) to cabbage (
Attachment strength of A. hydrophila cells to produce surfaces
The attachment strength (S R) of A. hydrophila cells to cabbage and lettuce surfaces increased with time (Fig. 4A, B). There was significant (p < 0.05) difference between S R of the biofilm strain TUB19 after 1 and 2 h on cabbage or lettuce surfaces; however, no significant (p > 0.05) differences between S R values of the other A. hydrophila strains were found. By extending the incubation time S R values were significantly increased. In general, the highest S R values were recorded by the biofilm strain TUB19 on both studied vegetable surfaces.
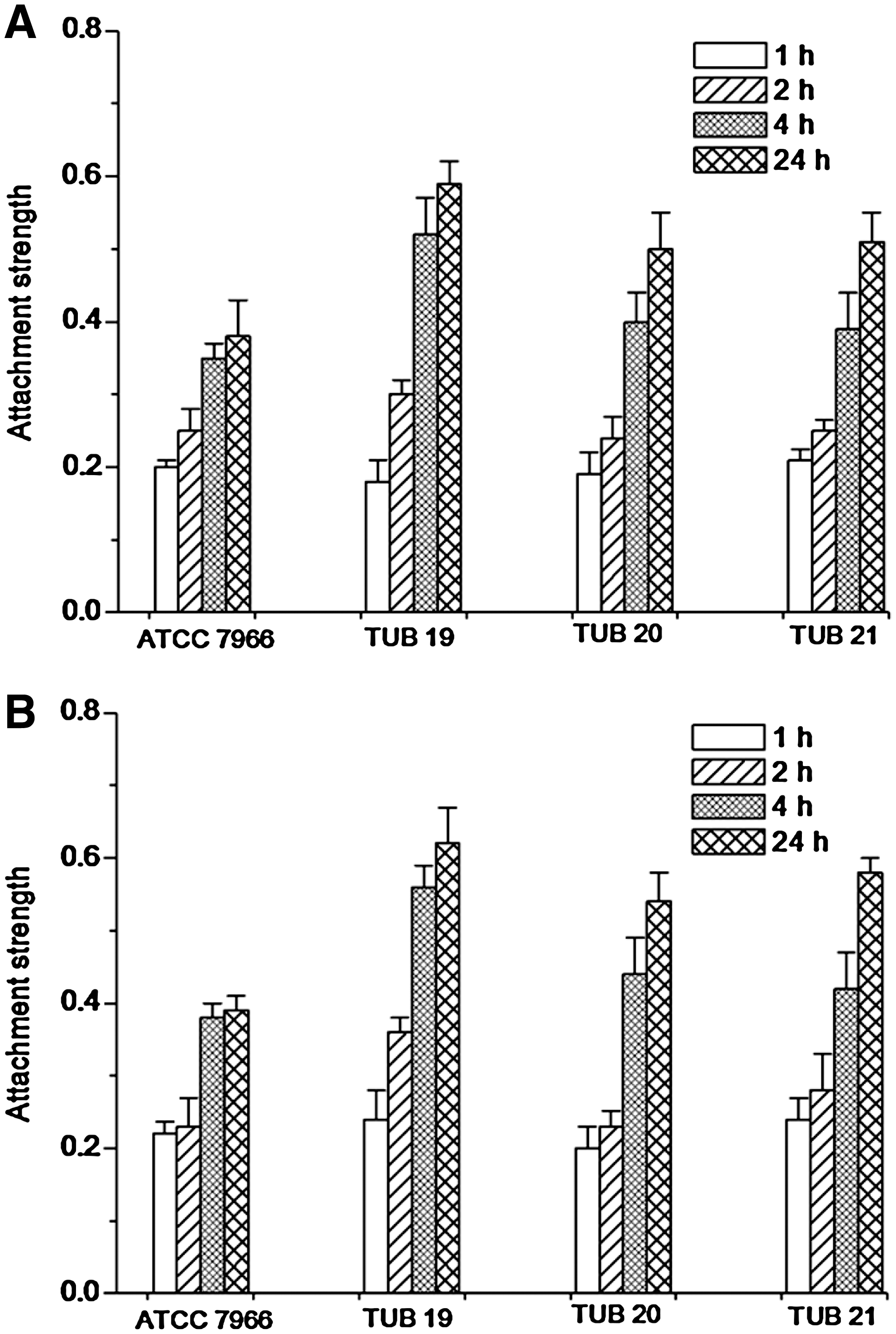
Attachment strength (S
R) of A. hydrophila strains (the reference strain ATCC 7966 and biofilm strains TUB19, TUB20, and TUB21) to cabbage (
Discussion
An improved understanding of the behavior of enteropathogenic bacteria, such as A. hydrophila on salad vegetables, should help prevent both the initial contamination events and microbial persistence up to the point-of-sale. Cell surface hydrophobicity of bacteria is a very important physicochemical feature, which has a great influence on the ability of bacteria to adhere to solid substratum (Ascencio et al., 1995; Gary et al., 2003). Bacterial cell surface hydrophobicity is affected by the molecular architecture of the outermost surface of bacteria. Therefore, the active attachment of bacterial cells to the substratum is affected by cell surface properties such as adhesion proteins, capsules, surface charge, flagella, and pili (Kumar and Anand, 1998). In the present study, the ability of biofilm strains was different from that of the reference strain. Also, changing the hydrophobicity of biofilm strains demonstrated in the present study may be due to exposure to the environment stress, where these strains have been isolated from biofilm samples collected from drinking water distribution networks. These differences in the cell surface hydrophobicity could be due to variation in the level of expression of cell surface proteins among strains as well as due to environmental conditions that could affect the expression of surface proteins (Ramiah et al., 2007). Presence of bacterial cells in the environment may lead to changes in cell surface properties influencing adhesion of the cells to the solid surfaces. This finding is in agreement with those mentioned by Khan (2009), who stated that, in some environmental strains, the tapY1 gene (encoding TypIV pili) played an important role in biofilm formation. Reduction in the in vitro biofilm formation of biofilm strains compared with the reference strain could be explained by low hydrophobicity measured. The hydrophobicity of bacterial cell surfaces can be of importance in many research areas, for example, biofouling (Chmielewski and Frank, 2003; Elhariry, 2008). Alternative factors that have been suggested to aid in vitro aeromonad adherence are motility, outer membrane proteins, the lipopolysaccharide (Aguilar et al., 1997) and the polar flagellum (Merino et al., 1997). In the microorganism adhesion to hydrocarbon test, removal of cells from the aqueous suspension depends on their adhesion to the hydrocarbon phase. Thus, this method is very sensitive to the amount of surface area created during mixing of the two liquid phases (Crow and Herndon, 1995; Elhariry, 2008).
Under starvation condition (TSB 1:20) all strains partially lost their ability to form biofilm on polystyrene surface. This may be due to reduction of mineral concentrations such as Mg2+ in diluted TSB. Previously, Merino et al. (2001) stated that MgtE (Mg2+ And Co2+ transport protein) and thus Mg2+ have a role in A. hydrophila related to their swarming ability and related consequences such as adherence and biofilm formation. Moreover, Asha et al. (2004) showed that 15 different proteins were repressed and 3 new proteins were expressed in biofilm of A. hydrophila under starvation conditions. Similar results were demonstrated for starved cells of Vibrio sp. lost many cellular proteins and synthesized new proteins called starvation proteins (Nelson et al., 1997). In other Gram-negative bacteria, Ngwai et al. (2006) stated that nutrient starvation led to reduction of biofilm formation by Salmonella. Stepanovic et al. (2004) hypothesized that stronger biofilm formation by Salmonella and Listeria in diluted TSB could be due to the induction of biofilm under starvation stress.
The mechanism by which the bacterial cells attach to a leaf surface is extremely complex and does not only rely on the topography of the surface (Ells and Hansen, 2006). For example, when materials possess pits and cavities on surfaces, as is observed in living surfaces, bacterial cells are more likely to be able to penetrate and embed themselves in these areas affording themselves partial if not complete protection from inactivation treatments. The mechanism is also considered to be a function of the physicochemical properties of both bacterium and plant surfaces (Hirano and Upper, 2000; Ukuku and Fett, 2002). A correlation between the negative surface charge and hydrophobicity of several bacterial pathogens and the strength of their adhesion to cantaloupe rind surfaces was previously studied by Ukuku and Fett (2002). They demonstrated also that the rind itself, with its coarse texture and irregular surface, promoted bacterial cell adhesion. The surface conditions of the produce may also play an important role in bacterial adhesion. Solomon and Sharma (2009) mentioned that bacteria with high hydrophobic characteristics may adhered to the cuticle, a hydrophobic layer on plant surfaces composed of fatty acids, polysaccharides, and waxes. Produce surfaces that are covered in hydrophobic waxy cuticles may allow hydrophobic A. hydrophila cells to adhere to the waxy cuticle. In agreement with the results obtained in the present work, Ells and Hansen (2006) reported that the relative strength of binding increased with prolonged exposure times (Fig. 4).
Results of the present work demonstrated that biofilm formation behavior by A. hydrophila cells on the polystyrene surface differed from that on the produce surface. This may be because plant surfaces are not homogeneous in nature, and they undergo dramatic changes during growth or after harvest and storage. Therefore, generalizations regarding the attachment, growth, and persistence of foodborne pathogens to synthetic surfaces may not hold true for plant surfaces. In contrast with this finding, Patel and Sharma (2010) indicated that in vitro biofilm formation by Salmonella might be correlated with attachment to some produce surfaces, even though in vitro biofilm formation and produce attachment studies were carried out at different temperatures. Therefore, further studies are required to improve our understanding the interaction between human microbial pathogens and surfaces of raw vegetables. Moreover, the use of microbial strains isolated from biofilm samples may be more useful for understanding of biofilm formation on green-leafy vegetables than the reference or laboratory strains.
Footnotes
Disclosure Statement
No competing financial interests exist.
Present address: Department of Food Science, Faculty of Agriculture, Ain Shams University, Cairo, Egypt.