
Abstract
We have investigated the distribution of integrons among 752 multidrug-resistant Salmonella isolates from human febrile and/or diarrheal patients during 1992–2007 and analyzed their genetic characteristics. Here, we report extensive integron analysis results within human isolates during the last 16 years. The gene or gene cassette(s) in the class 1 integrons found in the isolates were dfrA7, dfrA12-orfF-aadA2, aadA2, bla PSE1, dfrA1-aadA1, dfrA17-aadA5, bla OXA1 -aadA1, aadB-aadA1, aadA22, aadA1, and aac6'Ib-bla OXA1 -aadA2. Class 2 integrons harboring dfrA1-sat2-aadA1 gene cassette were also found in four isolates. Twenty-nine isolates including one Salmonella Schleissheim isolate had two integrons harboring aadA2 and bla PSE1 in their variable regions of 1.0 and 1.2 kb amplicons, respectively, which have been also found in Salmonella genomic island 1 (SGI1) of multidrug-resistant Salmonella Typhimurium DT104. The presence of SGI1 in Salmonella Schleissheim isolate was proved by SGI1-specific polymerase chain reaction. We first report a Salmonella Schleissheim having SGI1, Salmonella Typhimurium and Salmonella Heidelberg having the class 2 integron with dfrA1-sat2-aadA1 cassettes, Salmonella London with the aac6′Ib-bla OXA1 -aadA2 gene cassette, Salmonella Chailey with the gene cassette of aadA22, and coexistence of two class 1 integrons carrying aadA22 and dfrA12-orfF-aadA2 in Salmonella Typhimurium.
Introduction
Integrons are a kind of movable genetic element and play a pivotal role in the spread of gene cassette(s) horizontally (Hall, 1997). Integron-encoded integrase, a site-specific recombinase, is able to capture some genes including the antibiotic resistance genes in the variable region of the integron to form gene cassettes (Hall and Stokes, 1993; Rowe-Magnus and Mazel, 2001). Under antibiotic selective pressure or even in the absence of selective pressure, integrons could capture resistance gene(s) and contribute to lateral gene transfer using conjugative plasmids and/or transposons as vehicles. These foreign DNA parts including various integron structures can represent up to 15% of a given bacterial genome (Ochman et al., 2000).
Integrons can be classified based on the sequence of the integrase genes (Partridge et al., 2009). The class 1 integron has been commonly detected in Salmonella enterica. This integron consists of 5′ and 3′ conserved sequences and a variable region between them. The class 2 integron is an associated Tn7 transposon and has been less detected in Salmonella spp. than class 1 integrons and carries three resistance gene cassettes, dfrA1, sat1, and aadA1 (Hansson et al., 2002; Mazel, 2006). Many variable gene cassette arrays in the class 1 integron have been found in Salmonella serovars (Rodriguez et al., 2006; Vo et al., 2006).
Salmonella is one of the most frequent pathogens in foodborne diseases in Korea (Chung et al., 2004). Although an increasing trend of Salmonella antimicrobial resistance has been reported (Choi et al., 2005), there has not been a concrete report describing the influence of the integron on the antimicrobial resistance of Salmonella in Korea.
Therefore, to acquire information about the prevalence of integron and its different structures in Salmonella serovars, we investigated and analyzed the distribution of integrons in multidrug-resistant (MDR) Salmonella serovars isolated from diarrheal cases in Korea.
Materials and Methods
Bacterial isolates
A total of 752 Salmonella isolates that were resistant to >3 antibiotics were selected out of 15,357 isolates obtained from febrile and/or diarrheal patients in the course of the surveillance system in Korea, 1992–2007. Korea National Institute of Health has run the national surveillance system for the diarrheal diseases and isolated enteric bacterial pathogens from the patients. The isolates were identified by culture methods and biochemical tests with API-20E (BioMeriux) and were serotyped using O- and H-antisera according to the Spicer-Edwards scheme (Difco Laboratories).
Screening and sequencing of the variable regions of class 1 and 2 integrons
All the MDR 752 Salmonella isolates were screened first by polymerase chain reaction (PCR) using hep35 and hep36 primers that are specific to class 1, 2, and 3 integrons, and then class 1- and 2-specific PCRs were performed using 5′ CS/3′ CS and hep74/hep51 primer pairs, respectively, with the isolates that showed amplicon bands from the first-round PCR screening. PCR conditions and experiments for the integron screening were carried out as described elsewhere (Levesque et al., 1995; White et al., 2001).
The template DNA for PCR amplification was prepared by boiling a full bacterial colony resuspended in 100 μL deionized distilled water for 5 min and taking supernatants following centrifugation at 13,000 rpm for 10 min. DNAs were amplified in a total 30 μL reaction volume containing dNTPs, PCR buffer, template DNA, primers, and ExTaq polymerase according to manufacturer's instruction (Takara). For the analysis of the variable regions of integrons, all PCR products were purified with QIAquick PCR products purification kit (Qiagen) and subjected to direct sequencing (Macrogen).
Identification of Salmonella genomic island 1 in Salmonella Schleissheim
To identify Salmonella genomic island 1 (SGI1), PCR was performed with specific primers to amplify floR (forward primer: 5′-ATG ACC ACC ACA CGC CCC GCG TGG-3′, reverse primer: 5′-CTA GAC GAC TGG CGA CTT CTC GGT GGC-3′), tet(G) genes (Ng et al., 1999) located in SGI1, and the boundaries of SGI1 and In104 located in SGI1 (Boyd et al., 2001, 2002; Carattoli et al., 2002). For the positive control and for comparison, WSU2380 Salmonella Typhimurium DT104 USA isolate and KJ13 Salmonella Typhimurium DT104 Korean isolate were used (Kim et al., 2004). All PCR products were analyzed by agarose gel electrophoresis. The gel was stained with ethidium bromide (5 μg/mL), visualized with ultraviolet, and photographed using a GelDoc XR system (BioRad).
Similarity search of the sequence data
A similarity search was performed using the BLAST program found on the NCBI BLAST homepage (
Results and Discussion
Distribution of class 1 and 2 integrons in various Salmonella serovars and gene cassette(s) in integrons
Among 752 MDR Salmonella isolates, 144 isolates (19.1%) were positive for the class 1 integron PCR and 4 isolates (0.5%) were positive for the class 2 integron PCR. Of the 144 Salmonella isolates showing class 1 integron PCR positive, the numbers of Salmonella Typhimurium, Salmonella Enteritidis, and Salmonella Typhi isolates were 96 (66.7%), 18 (12.5%), and 10 (6.9%), respectively. The other serovars of Salmonella were Anatum, Montevideo, Paratyphi B, Braenderup (2 isolates, each), Chailey, Corvalis, Derby, Heidelberg, Lexington, Lomita, London, Muenchen, Rissen, S. salamae, Schleissheim, and the untypable (1 isolate, each) (Table 1).
UT, untypable serovar.
The integron PCR results from the class 1- and 2-specific primers were differentiated into 14 profiles by their sizes and the band patterns (Table 1). The cassette genes in class 1 integrons were the genes encoding dihydrofolate reductase A (dfrA1, dfrA7, dfrA12, dfrA17), aminoglycoside adenyltransferase A and B (aadA1, aadA2, aadA5, aadA22, aadB), β-lactamase (bla PSE1, bla OXA1), aminoglycoside 6′-N-acetyltransferase (aac6′Ib), and hypothetical protein F (orfF). The 0.7-kb PCR product was the most common integron gene cassette (42 isolates, 28.4%) in this study, which encodes a dfrA7 between 5′ CS and 3′ CS, conferring resistance to trimethoprim.
All of nine MDR Salmonella Typhi isolates, only one isolate contained this dfrA7 cassette in class 1 integron, which had been reported in MDR Salmonella Typhi isolated in Korea and Nepal (Lee et al., 2004; Tamang et al., 2007). This gene cassette was also found in 25 Salmonella Typhimurium isolates. The other serovars and subspecies having dfrA7 cassette in class 1 integron were Salmonella Enteritidis (six isolates), Salmonella Montevideo (one isolate), and S. salamae (one isolate). This result indicates that class 1 integron harboring dfrA7 single cassette might have been transferred horizontally to various Salmonella serovars from 1999 or acquired from other sources.
The second-most widespread integron was the class 1 integron harboring three gene cassettes, dfrA12-orfF-aadA2 (41 isolates, 27.7%), conferring resistance to trimethoprim and aminoglycosides. This integron structure was identified in serovars Salmonella Anatum (2 isolates), Salmonella Enteritidis (5 isolates), Salmonella Lexington (1 isolate), Salmonella Lomita (1 isolate), Salmonella Typhimurium (31 isolates), and the untypable (1 isolate). An integron with this gene cassette was also reported as the most frequent in Escherichia coli from humans, poultry, and swine from 2001 to 2003 in Korea (Kang et al., 2005). So, this class 1 integron with dfrA12-orfF-aadA2 cassettes has been circulated widely among Salmonella serovars as well as E. coli.
A type of class 1 integron with dfrA17-aadA5 was identified in a Salmonella Enteritidis isolate and a Salmonella Typhimurium isolate, respectively. This type of integron had been the most prevalent structures in clinical isolates of E. coli in Korea. However, class 1 integron containing dfrA1-aadA1, which was identified in three isolates in this study, had been rare in both serovars as well as in other enterobacteriaceae in Korea (Yu et al., 2003; Kang et al., 2005).
Two class 1 integrons containing bla OXA1 were each detected in a single serovar. One was an integron containing bla OXA1 -aadA1 gene cassettes in Salmonella Muenchen and the other was an integron containing aac6′Ib-bla OXA1 -aadA2 arrays in Salmonella London. The aac6′Ib gene encodes an aminoglycoside, 6′-N-acetyltransferase, conferring resistance to aminoglycosides. In America, bla OXA30 -aadA2 on a class 1 integron was found in MDR Salmonella Muenchen porcine isolates (Gebreyes and Thakur, 2005). These two integron structures were very similar between the MDR Salmonella Muenchen isolates because bla OXA30 is bla OXA1 (Boyd and Mulvey, 2006). To our knowledge, the aac6′Ib-bla OXA1 -aadA2 gene cassettes in Salmonella spp. is the first report in the world. The gene cassettes have been also found in a plasmid of a Klebsiella pneumoniae isolate (GenBank accession number: AF227505).
Among the minor class 1 integrons, aadA1, aadA22, and aadB-aadA1 gene cassettes were identified in serovars Salmonella Corvallis, Salmonella Chailey, and Salmonella Typhimurium, respectively. A Salmonella Typhimurium isolate harboring two class 1 integrons, aadA22 and dfrA12-orfF-aadA2 together, was also found. The aadA22 that confers high-level resistance to streptomycin and spectinomycin has been reported very recently in Salmonella Choleraesuis (Lee et al., 2009). Interestingly, the aadA22-carrying pathogen was Salmonella Chailey, a very rare Salmonella serovar in Korea as well as in the world. Taken together, aadA22 in Salmonella Chailey and coexistence of two class 1 integrons carrying aadA22 and dfrA12-orfF-aadA2, respectively, in Salmonella Typhimurium are the first reports regarding the distribution of integrons.
A type of class 2 integron with dfrA1-sat2-aadA1 cassettes was identified in two isolates of Salmonella Typhimurium, one isolate of Salmonella Enteritidis, and one isolate of Salmonella Heidelberg. sat2 confers resistance to streptothricin. Class 2 integrons were identified and reported in Salmonella Enteritidis (Ahmed et al., 2009), Salmonella Paratyphi A (Miko et al., 2003), Salmonella Virchow, Salmonella Grumpensis, Salmonella Worthington, and Salmonella Panama (Rodriguez et al., 2006). This is the first report that Salmonella Typhimurium and Salmonella Heidelberg have the class 2 integron with dfrA1-sat2-aadA1 cassettes. The five genetic structures of the predominant integrons in this study are shown in Figure 1.
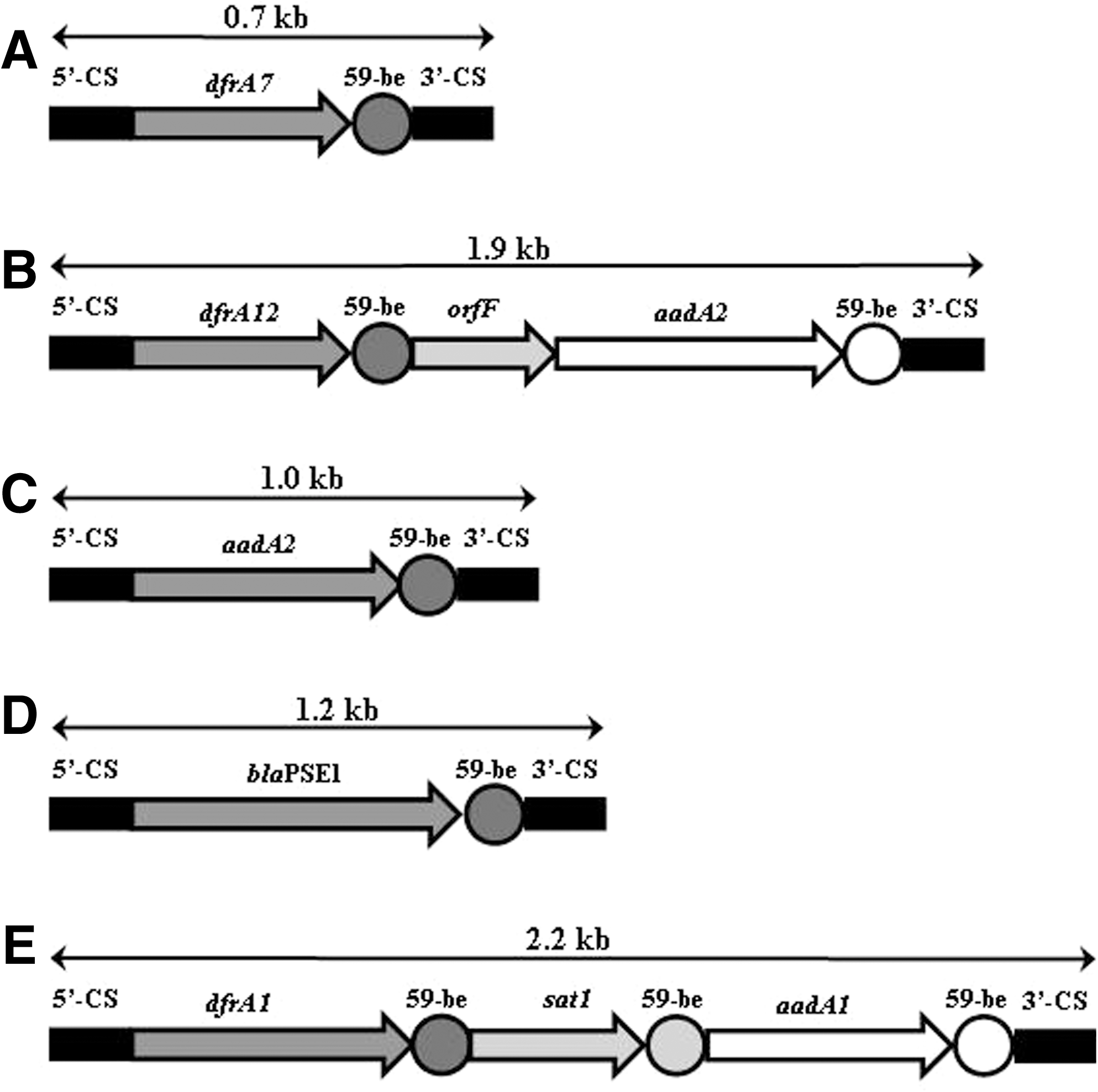
The five genetic structures of the predominant class 1
SGI1 in Salmonella Schleissheim
SGI1 has been initially found in MDR Salmonella Typhimurium DT104. The size of SGI1 is ∼43 kb, which is located between thdF and int2. However, SGI1 is located between chromosomal genes thdF and yidY in other Salmonella serovars. The SGI1 harbored an antibiotic resistance gene cluster in its 3′ region flanked by two class 1 integrons. Its size was ∼13 kb in length and it is called “In104” (Briggs and Fratamico, 1999; Boyd et al., 2001). Recently, SGI1 and its derivatives have been found in the genome of MDR Salmonella Typhimurium DT1, DT12, DT120, and U302 (Boyd et al., 2002; Carattoli et al., 2002; Lawson et al., 2002) and other Salmonella serovars Agona, Paratyphi B, Albany, Meleagridis, Newport, Cerro, Derby, Dusseldorf, Infantis, Kiambu, and Emek (Cloeckaert et al., 2000; Boyd et al., 2002; Meunier et al., 2002; Doublet et al., 2003, 2004a, 2004b, 2004c; Ebner et al., 2004; Levings et al., 2005).
In the present work, aadA2 and bla PSE1, which are parts of In104 located in SGI1, were identified independently or together in various serovars (Table 1). Salmonella isolates having both integrons showed more prevalence (19.6%) than the isolates having a single integron. It was thought that the isolates having both integrons had In104 region located in SGI1 like Salmonella Typhimurium DT104 (Levings et al., 2005). In fact, the number of Salmonella Typhimurium isolates having both integrons was 26, indicating that Salmonella Typhimurium was the major serovar among other serovars having integrons with aadA2 and bla PSE1 in this study. SGI1 had been found and characterized first in serovar Typhimurium DT104 (Boyd et al., 2001). The other serovars were Salmonella Derby, Salmonella Paratyphi B, and Salmonella Schleissheim. Except Salmonella Schleissheim, the presence of SGI1 in these serovars genome had been reported.
The Salmonella Schleissheim isolate (KJ18919) was screened by PCR for the presence of In104 and SGI1 with MDR Salmonella Typhimurium DT104 isolates WSU2380 and KJ13 as positive controls as written in the Materials and Methods section. WSU2380 and KJ13 isolates have an indistinguishable XbaI pulsed-field gel electrophoresis pattern and were isolated from an American cattle and a Korean patient, respectively (Kim et al., 2004).
Both DT104 isolates and the Salmonella Schleissheim isolate had both ends of In104 in SGI1. The Salmonella Schleissheim isolate possessed both ends of SGI1. However, both DT104 isolates did not show positive PCR result for right junction of SGI1, consistent with data from other DT104-related studies, because Salmonella Typhimurium DT104 with SGI1 contains a retron element that disrupts the right junction of SGI1 (Levings et al., 2005). To determine if, like DT104, the Salmonella Schleissheim isolate possesses floR and tet(G) genes, specific primers were designed and used. PCR results showed expected products of the sizes of the floR and tet(G) genes (Table 2).
SGI, Salmonella genomic island; LJ, left junction; RJ, right junction; +, PCR positive; −, PCR negative.
Changes in the genetic structures of integrons during 16 years
In this study, we were able to identify the distribution and genetic structures of integrons among 752 isolates of MDR Salmonella from human patients from 1992 to 2007. During that time, integrons in the MDR isolates with dfrA7 or dfrA12-orfF-aadA2 cassettes have been circulated until recently for 6–7 years but dfrA1-sat2-aadA1 genetic structure has not been found since 2001. The integrons harboring aadA1 or aadA22 were found at the end of the study period in 2007. The coexistence of an integron with aadA2 and an integron with bla PSE1 in an isolate that has possibly SGI1 has been epidemic for 12 years and detected in various Salmonella serovars since 1994 (Table 1). According to these results, an integron-related genetic epidemiological data for 16 years in Korea were presented.
Footnotes
Acknowledgments
The authors thank Ms. Esther Lee for her help with PCR experiments. This study was supported by an intramural grant from the Korea Centers for Disease Control and Prevention.
Disclosure Statement
No competing financial interests exist.