
Abstract
Campylobacter is one of the most common foodborne pathogens causing acute gastroenteritis in humans. Erythromycin, a macrolide antibiotic, is the first-choice treatment for Campylobacter infections, and failure to eradicate Campylobacter is usually due to macrolide resistance. The most important mechanism responsible for macrolide resistance in Campylobacter is mediated by point mutations at position 2074 or 2075 in the peptidyl-transferase region of domain V of the 23S rRNA gene. In this study, the minimum inhibitory concentrations of 58 Campylobacter isolates (C. jejuni: n = 37; C. coli: n = 21) obtained from chickens were measured by agar dilution. Isolates were subjected to both pyrosequencing and Sanger sequencing methods to detect the 2074 and 2075 point mutations and evaluate the efficacy of the pyrosequencing method. The A2075G mutation was found to be the predominant mutation associated with erythromycin resistance. Compared with traditional methods, pyrosequencing is a novel, rapid, low-cost, and quantitative technology for detecting erythromycin resistance in Campylobacter.
Introduction
Bacteria have evolved several mechanisms for macrolide resistance, including target modification by the existence of point mutations or methylation of the 23S rRNA gene, hydrolysis of the drug, and efflux pumps (Aarestrup and Engberg, 2001). In Campylobacter, point mutations at positions 2074 or 2075 in the 23S rRNA peptidyl-transferase gene are the main cause of macrolide resistance. The most prevalent mutations at these positions are adenine-to-cytosine and adenine-to-guanine (Gibreel et al., 2005; Gibreel and Taylor, 2006; Alfredson and Korolik, 2007). Further, recent studies revealed that at least two of the three 23S rRNA copies must be mutated to confer macrolide resistance (Gibreel et al., 2005). To test the susceptibility profiles of Campylobacter strains, there are several phenotype-based methods including agar dilution, broth dilution, and the E-test (CLSI, 2008). Several molecular detection techniques, such as the polymerase chain reaction (PCR) line probe assay (Niwa et al., 2001, 2003), PCR–restriction fragment length polymorphism (Vacher et al., 2003), mismatch amplification mutation assay–PCR (Payot et al., 2004; Alonso et al., 2005), SYBR green real-time PCR (Vacher et al., 2005), and TaqMan real-time PCR (Hao et al., 2010), have been used to detect macrolide resistance mutations in the 23S rRNA gene of Campylobacter. Most of those methods are qualitative assays that can be used to distinguish between isolates with homozygous alleles and heterozygous alleles, but cannot be used to detect the accurate number or proportion of alleles with mutated genes. As mentioned previously, macrolide resistance is dependent on the number of mutated copies of the gene, so it is necessary to develop a quantitative assay to identify the proportion of mutated alleles. In pyrosequencing, the peak heights of the pyrograms reflect the number of nucleotides incorporated according to the template. Pyrosequencing is a rapid, low-cost method to detect single-nucleotide polymorphisms (SNPs) of point mutations associated with antimicrobial resistance in clinical isolates (Moder et al., 2007; Gorgani et al., 2009).
In the present study, we developed a rapid and quantitative assay to detect point mutations at position 2074 or 2075 of the 23S rRNA gene (rDNA), which play a key role in erythromycin resistance in C. jejuni and C. coli isolates. This assay can be potentially used to acquire important information regarding Campylobacter macrolide resistance in China.
Materials and Methods
Bacterial isolates and erythromycin susceptibility testing
In total, 58 Campylobacter isolates (including 37 C. jejuni isolates and 21 C. coli isolates) were obtained from chicken cecal contents and identified by multiplex PCR. Sampling, isolation, and identification procedures were performed as previously described (Chen et al., 2010). The minimum inhibitory concentrations (MICs) of these isolates against erythromycin (provided by the China Institute of Veterinary Drug Control, Beijing, China) were determined by agar dilution with 5% defibrinated sheep blood according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2008). C. jejuni strain ATCC 33560™ (American Type Culture Collection) was used as a control strain.
Genomic DNA extraction and PCR amplification of the 23S rRNA gene
Genomic DNA of 58 Campylobacter isolates was extracted using the TIANamp Bacteria DNA Kit (TIANGEN) and used as a PCR template for the following studies. Previously reported primers against the three copies of the 23S rRNA V domain were used in PCR amplification (Vacher et al., 2003), as listed in Table 1. PCR mixtures consisted of both primers (23-F and 23-R) at a concentration of 1 μM, 10 × ExTaq buffer (Mg2+ plus; TaKaRa), 250 μM of dNTPs (TaKaRa), 1 U of ExTaq polymerase (TaKaRa), 150 ng of template DNA, and sterilized distilled water up to 50 μL. The reactions were performed in a Veriti 96-well Thermal Cycler (Applied Biosystems) and the cycling parameters were 95°C for 5 min, followed by 35 cycles of 95°C for 1 min, 55°C for 40 sec, and 72°C for 1 min, followed by a final extension step at 72°C for 10 min. Amplification was confirmed by 1% (w/v) agarose gel electrophoresis and a 508-bp product was expected.
Numbering of the nucleotide position follows the NCTC11168 (GenBank accession number is AL11168.1).
PCR, polymerase chain reaction.
Sanger sequencing for part of 23S rRNA gene
PCR products were purified using the TIANgel Midi Purification Kit (TIANGEN) according to the manufacturer's instructions. Forward and reverse primers to the 23S rRNA gene (Table 1) were used as sequencing primers. Sequencing was performed by Invitrogen.
Pyrosequencing protocol
PCR amplification
The PCR primers and pyrosequencing primers to the 23S rRNA gene were based on the sequence of C. jejuni strain NCTC11168 (GenBank accession number: AL11168.1) and designed using the PSQ assay design program (Biotage). The reverse primer was tagged with biotin at the 5′ end. All primers are listed in Table 1. All PCR amplifications were performed in a final volume of 50 μL containing 0.2 μM of each primer (Pyro-F and Pyro-R), 5 μL of 10 × ExTaq buffer (TaKaRa), 2 mM Mg2+, 200 μM of each dNTP (dATP, dTTP, dGTP, and dCTP) (TaKaRa), 1 U ExTaq polymerase (TaKaRa), 150 ng template DNA, and sterilized distilled water. The PCRs were performed in a Veriti 96-well Thermal Cycler (Applied Biosystems) and cycling parameters were 95°C for 10 min, followed by 45 cycles of 95°C for 1 min, 60°C for 45 sec, and 72°C for 45 sec and a final extension at 72°C for 10 min. Amplification products were checked by 1.2% (w/v) agarose gel electrophoresis and a 227-bp product was expected.
Sample preparation for pyrosequencing
Single-stranded DNA (ssDNA) templates for pyrosequencing were obtained according to the manufacturer's instructions (Biotage). Briefly, 40 μL of binding buffer (pH 7.6; 10 mM Tris-HCl, 2 M NaCl, 1 mM ethylenediaminetetraacetic acid, 0.1% Tween-20) and 2 μL streptavidin-coated sepharose beads (GE Healthcare) were added to the biotinylated amplicon and then agitated at a speed of 1300–1400 rpm for 10 min to allow binding of the biotinylated amplicon. The amplicon was washed with 70% ethanol and denatured with 0.2 M NaOH to release ssDNA. The ssDNA was washed with 10 mM Tris-acetate (pH 7.6) and transferred to a 96-well microtiter plate containing 40 μL of annealing buffer (pH 7.6; 20 mM Tris-acetate, 2 mM Mg-acetate) and 0.3 μM of the sequencing primer. The plate was heated to 80°C for 2 min and then cooled to room temperature.
Pyrosequencing
Pyrosequencing was performed with PyroGold reagent (Biotage) on a PSQ 96MA instrument (Biotage). All subsequent steps were conducted according to the manufacturer's instructions. The dispensation order AGAC3 (GATC) was designed for confirming the amount of mutated alleles at positions 2074 and 2075 (corresponding to positions 2058 and 2059 in Escherichia coli) using the SNP mode of the PSQ 96MA software.
Results
The MICs of erythromycin for the 58 isolates were determined by the agar dilution method. According to the CLSI recommended breakpoint (CLSI, 2008) for the C. jejuni isolates, 11 showed high-level resistance to erythromycin (MIC ≥256 μg/mL), 22 showed low-level resistance to erythromycin (MIC 32–128 μg/mL), and 4 were susceptible to erythromycin (MIC ≤32 μg/mL). All 21 of the C. coli isolates tested showed high-level resistance to erythromycin.
In this study, we used pyrosequencing technology to identify mutations in the 23S rRNA V domain of a small number of Campylobacter isolates. C. jejuni strain ATCC 33560 was used as a negative control. Sanger sequencing confirmed the pyrosequencing results. The predominant mutation associated with erythromycin resistance is shown in Figure 1. The wild-type sequence obtained by pyrosequencing showed a high peak of A (adenine) at positions 2074–2076, which corresponds to the incorporation of three adenine nucleotides (Fig. 1a), whereas the pyrogram of the mutant sequences conferring erythromycin resistance read AGA (A2075G mutation) (Fig. 1b). The SNP mode (A: 1.4%; G: 98.6%) indicated A2075G mutations in all three copies of 23S rRNA gene.
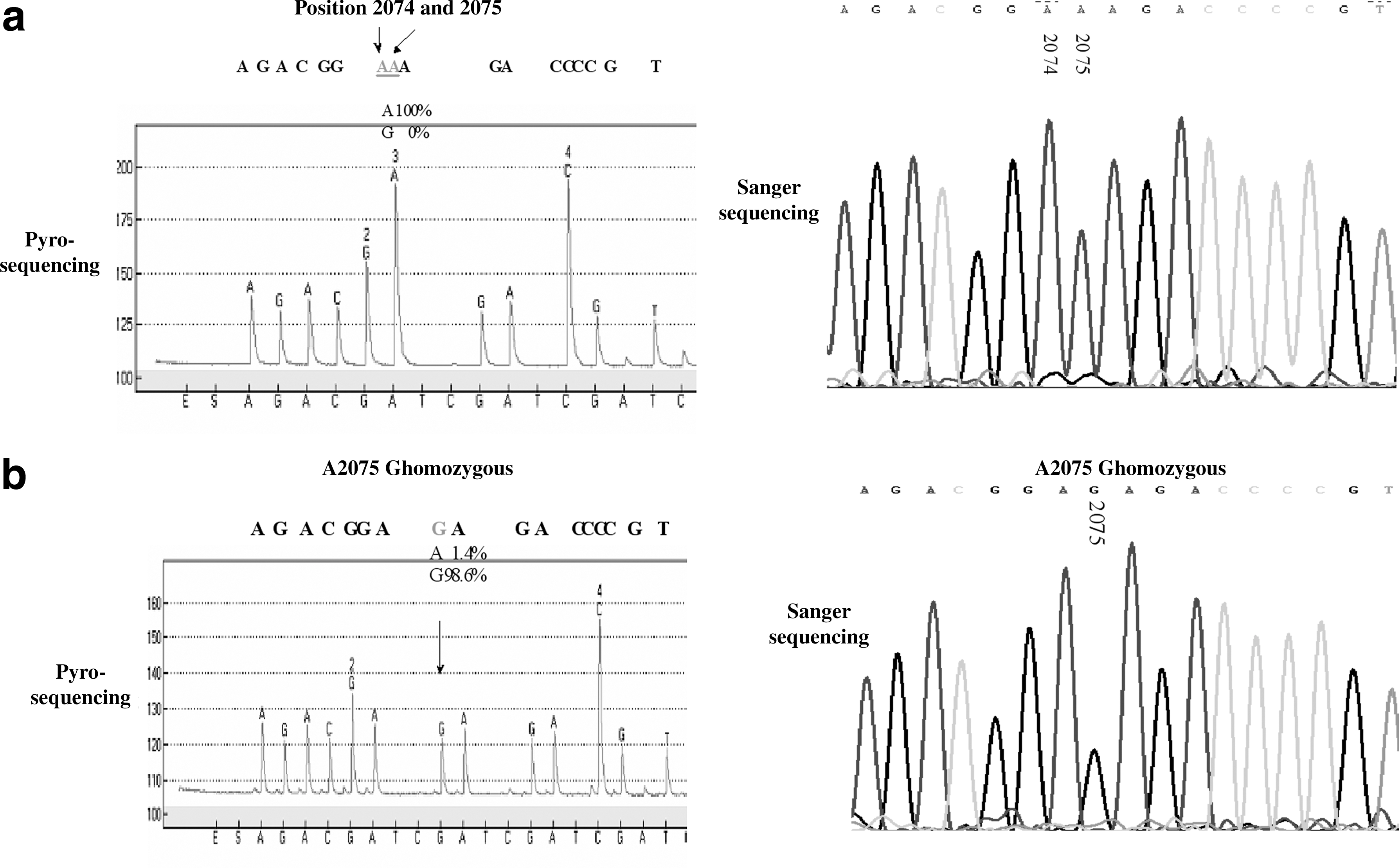
Pyrosequencing and Sanger sequencing of the 2074 and 2075 positions of the 23S rRNA gene of Campylobacter jejuni and Campylobacter coli.
The correlation between mutations revealed by pyrosequencing and the erythromycin resistance profile of Campylobacter isolates is shown in Table 2. Generally, for the 58 isolates tested, 22 of the low-level erythromycin-resistant Campylobacter isolates harbored wild-type sequences, whereas most of the high-level erythromycin-resistant Campylobacter isolates (7 C. jejuni isolates and 21 C. coli isolates) harbored three copies of the mutation A2075G in the 23S rRNA gene, as shown by pyrosequencing. Other genotypes associated with erythromycin resistance were not identified in this study. All erythromycin-sensitive isolates and four of the highly erythromycin-resistant C. jejuni isolates showed no detectable mutations.
ND, not detected.
Discussion
The conventional antimicrobial susceptibility testing methods for Campylobacter spp. involve incubation of the organisms, and are therefore time-consuming to perform, and require specific expertise. Therefore, rapid and reliable methods are needed for the detection of erythromycin resistance caused by point mutations in these organisms. In recent years, PCR-based methods, such as PCR–restriction fragment length polymorphism (Vacher et al., 2003), mismatch amplification mutation assay–PCR (Alonso et al., 2005), and SYBR green real-time PCR (Vacher et al., 2005), have been used to detect mutations in the 23S rRNA of Campylobacter spp. Compared with these methods, pyrosequencing, as described in the present study, has the benefits of being inexpensive and simple to perform. Pyrosequencing is also a rapid method, taking <1 h to perform, and as many as 96 samples can be processed at one time. A further major benefit of this method is that it can be used as a quantitative assay, detecting the proportion of incorporated nucleotides.
Previous studies have investigated the relationship between point mutations and the phenotype of resistance to erythromycin in Campylobacter spp. High-level erythromycin resistance in Campylobacter spp. is mainly due to mutations in domain V of the 23S rRNA at positions 2074 and 2075, and the A2075G mutation is more frequently observed than the A2074G mutation (Gibreel and Taylor, 2006). Our results confirmed A2075G as the predominant mutation associated with erythromycin resistance in C. jejuni and C. coli strains, and this mutation occurred in all alleles. No mutation at position 2074 was detected in our study. A recent study highlighted the fact that the 2074 mutations (A2074C or A2074G) are only observed in vitro (Caldwell et al., 2008). They reported that with respect to high-level erythromycin resistance, all mutants selected in vivo displayed an A2074G mutation, whereas all mutants selected in vitro displayed an A2074C mutation in the 23S rRNA gene. This may explain why only A2075G changes, and no 2074 mutations, were identified in our study because all of our strains were isolated from chickens. Regarding the four isolates lacking mutations in their 23S rRNA genes but exhibiting erythromycin resistance, other resistance mechanisms may exist in these isolates. It has been proposed that the efflux pump CmeABC (Lin et al., 2002; Pumbwe and Piddock, 2002; Cagliero et al., 2005; Mamelli et al., 2005) and/or mutations in ribosomal proteins L4 and L22 (Cagliero et al., 2006; Caldwell et al., 2008) may be responsible for erythromycin resistance in these Campylobacter isolates.
In conclusion, pyrosequencing is a novel approach to analyze the genetic basis for antimicrobial resistance. This study clearly shows that pyrosequencing is a rapid, reliable, and cost-effective technique to determine the patterns of macrolide resistance among Campylobacter strains.
Footnotes
Acknowledgments
The authors thank Ms. Rachel Sippy (Iowa State University) for revision of this manuscript. This study was supported by grants from the Program for Chang Jiang Scholars and the Innovative Research Team at the University of China (No. IRT0866) and the Exclusive Research Fund for Public Welfare from the Ministry of Agriculture of the People's Republic of China (No. 200903055).
Disclosure Statement
No competing financial interests exist.