
Abstract
Twenty Pseudomonas aeruginosa isolates were recovered from imported frozen raw shrimp sold in the United States. Isolates were tested for antimicrobial susceptibility to quinolones and analyzed for mutations in quinolone resistance-determining regions, presence of type III secretion system genes, and genetic relatedness using pulsed-field gel electrophoresis. All isolates were resistant to nalidixic acid. Polymerase chain reaction assays detected exoS, exoT, exoU, and exoY among isolates. Eight unique pulsed-field gel electrophoresis clusters were generated. Mutations were found in gyrA at codon 83 (Ile to Thr) and in parC at codon 87 (Leu to Ser). Together, these findings reveal that imported shrimp may harbor virulent and quinolone-resistant strains of P. aeruginosa.
Introduction
The pathogenesis of P. aeruginosa is due to the production of several extracellular virulence factors, ExoS, ExoT, ExoU, and ExoY, encoded by a type III secretion system. Expression of ExoU/ExoT or ExoS/ExoT causes damage to the mucosal barrier, inhibits innate immunity, and prevents wound healing (Engel and Balachandran, 2009). The presence of exoU and fluoroquinolone resistance have been suggested as co-selected traits resulting in highly virulent and resistant strains of P. aeruginosa (Wong-Beringer et al., 2008), which warrants further studies on virulence genes and fluoroquinolone resistance.
In developing countries, antibiotics are widely used by the shrimp aquaculture industry for both prophylaxis and treatment of disease outbreaks in shrimp (Holmstrom et al., 2003). Further, multidrug-resistant bacteria have been isolated from shrimp (Le et al., 2005). This study describes the isolation, characterization, and detection of type III secretion system genes and mutations in quinolone-resistance–determining regions present in quinolone-resistant P. aeruginosa from imported shrimp.
Materials and Methods
Twenty isolates of P. aeruginosa were isolated from a total of 10 samples of farm-raised, frozen, whole raw shrimp (Penaeus spp.) imported from India and purchased from one retail store. One gram of shrimp was taken from three individual bags and homogenized in a stomacher with 10 mL of LB broth supplemented with 10 μg/mL of nalidixic acid. The homogenate was enriched overnight at 37°C and 100–200 μL was subcultured onto TSAII plates. Isolates were confirmed as P. aeruginosa using VITEK2 Gram-negative identification cards.
Antimicrobial minimum inhibitory concentrations were determined using the Sensititre Automated Antimicrobial Susceptibility System according to manufacturer's instructions and interpreted with Clinical Laboratory Standards Institute (CLSI) criteria (2010).
Pulsed-field gel electrophoresis (PFGE) analysis was performed using the Centers for Disease Control and Prevention procedure for Salmonella (
Polymerase chain reaction (PCR) assays were performed in a final volume of 25 μL, using 23 μL of SuperMix Hi Fidelity (Invitrogen, Carlsbad, CA), 50 pmol of each forward and reverse primer, and 1 μL of genomic DNA as template. The amplification of gyrA (∼400 bp), gyrB (∼370 bp), parC (∼180 bp), and parE (∼600 bp) genes was performed with previously published primers and amplification conditions (Mouneimne et al., 1999; Akasaka et al., 2001). Positive PCR products were purified with a Qiaquick PCR Purification Kit (Qiagen, Valencia, CA) and sequenced (MWG Biotech, Huntsville, AL). DNA sequences were analyzed by comparison with published sequences using the Pseudomonas Genome Database BLAST (
Results and Discussion
PFGE analysis of SpeI restriction digests of DNA from 20 P. aeruginosa isolates produced eight distinct profiles, each containing 14–20 restriction fragments. Profiles were clustered based on patterns with a genetic similarity of 95% or more (Fig. 1).
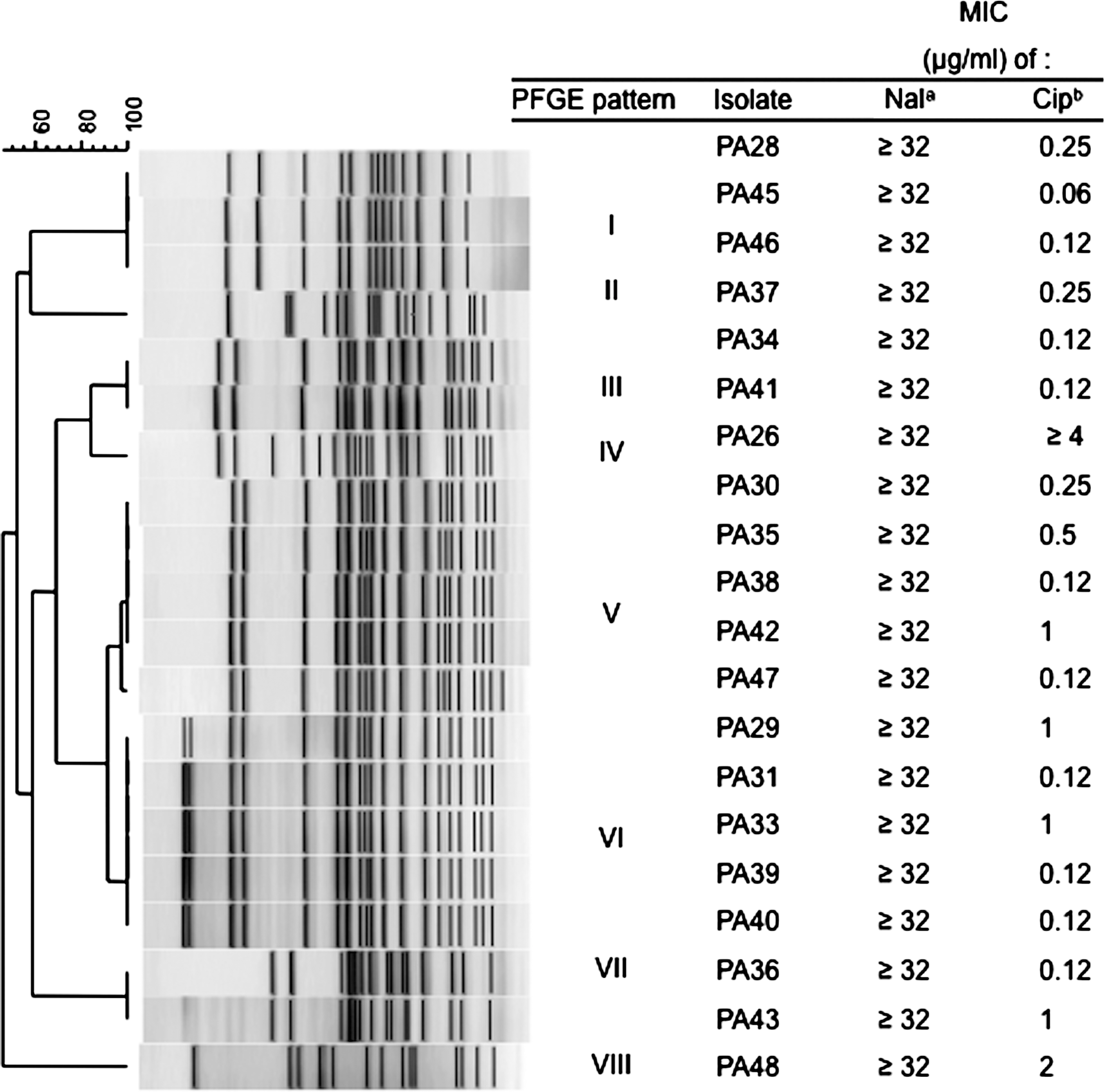
Unweighted pair group means with arithmetic averages analysis of PFGE profiles of Pseudomonas aeruginosa isolates from imported shrimp digested with SpeI (Clusters I–VIII). Fluoroquinolone MICs for P. aeruginosa isolates from imported shrimp. aResistance breakpoint (μg/mL) for Nal (nalidixic acid) according to CLSI standards (CLSI, 2010) is ≥32. bResistance breakpoint (μg/mL) for Cip (ciprofloxacin) according to CLSI standards (CLSI, 2010) is ≥4. MIC, minimum inhibitory concentration; PFGE, pulsed-field gel electrophoresis.
All isolates were resistant to nalidixic acid (≥32 μg/mL). PA26 demonstrated resistance to ciprofloxacin (≥4 μg/mL) and PA48 exhibited intermediate resistance to ciprofloxacin (2 μg/mL). Minimum inhibitory concentration values determined for nalidixic acid and ciprofloxacin corresponding to P. aeruginosa isolates grouped by PFGE patterns are displayed in Figure 1. Isolates also displayed resistance to other antibiotics, including β-lactams, aminoglycosides, and tetracycline.
Amino acid replacement in the quinolone resistance determining region of gyrA (Thr-83→Ile) was found in all 20 isolates. This finding was in accordance with previous reports on clinical isolates of P. aeruginosa (Mouneimne et al., 1999). The amino acid sequences in the quinolone resistance determining region of parC showed replacement of Ser-87 to Leu in all isolates. Notably, all isolates possessed alterations in gyrA and parC, similar to predominant alterations in clinical isolates (Akasaka et al., 2001). No mutations in gyrB nor parE were found, which agreed with previous studies reporting that these mutations are rare in P. aeruginosa (Piddock et al., 1999).
The identification of P. aeruginosa type III effector proteins, ExoS, ExoT, ExoU, and ExoY, was investigated. The exoS gene was amplified from 17 of 20 isolates. A previous study found variability in the presence of exoS among clinical samples (Feltman et al., 2001), which concurs with our findings that not all P. aeruginosa possess the exoS sequence. exoT was amplified from 16 of 20 isolates. This differed from Feltman et al. (2001), who reported exoT as an invariable trait present in all P. aeruginosa isolates. The discrepancy may be due to difficulty in discriminating between exoS and exoT sequences, since they share 80% of genetic identity or in sources of isolates examined. The exoU gene was amplified from 7 of 20 isolates. Similar to findings in clinical and environmental isolates of P. aeruginosa, exoU was found in a minority of the isolates (Feltman et al., 2001). The exoY gene was present in the majority of isolates, 17 of 20 isolates, but the biological function of ExoY remains unclear (Engel and Balachandran, 2009).
In the present study, the identification of chromosomal mutations in the gyrA and parC genes of quinolone-resistant P. aeruginosa isolates from shrimp and type III secretion system genes help supports the concept that P. aeruginosa isolated from shrimp may harbor virulence and quinolone-resistant determinants.
Footnotes
Acknowledgments
We thank Steve Yan and John B. Sutherland for critical review of the manuscript. This work was supported by the National Center for Toxicological Research, U.S. Food and Drug Administration. Q.T.T. and K.T.N. were supported by the FDA Commissioner's Fellowship Program. Views presented here do not necessarily reflect those of the FDA.
Disclosure Statement
No competing financial interests exist.