
Abstract
Cronobacter sakazakii is an emerging foodborne pathogen transmitted exclusively through contaminated infant formula (IFM), and associated with life-threatening infections in infants. C. sakazakii has the ability to tolerate a variety of environmental stress conditions, including heat stress, acidity, high osmotic pressure, and desiccation. In this study, we investigated the efficacy of a subinhibitory concentration (750 μM) of trans-cinnamaldehyde (TC), an ingredient in cinnamon, for reducing C. sakazakii's tolerance to these environmental stresses. Three strains of TC-treated C. sakazakii were separately subjected to high temperature (50°C, 55°C, and 60°C), acidic pH (3.3), high osmotic pressure (aw 0.81), and desiccation. TC (750 μM) substantially (p < 0.05) compromised stress tolerance of C. sakazakii compared to C. sakazakii cells not exposed to TC. Real-time quantitative polymerase chain reaction results revealed that TC significantly (p < 0.05) downregulated C. sakazakii genes critical for stress tolerance and survival, including rpoS, chaperonins, phoP/Q, outer membrane porins, and osmolyte transporter genes. The efficacy of TC in reducing C. sakazakii stress tolerance underscores its potential use for controlling the pathogen by increasing its susceptibility to commonly applied hurdles in food processing.
Introduction
Bacteria in the environment are exposed to a variety of stresses that include fluctuations in temperature, nutrient and water availability, pH, and the presence of toxic molecules, all of which can compromise their survival. C. sakazakii encounters these stresses in the environment, during its association with the IFM and within the host after infection. The ability of C. sakazakii to resist adverse environmental stresses is one of the critical factors responsible for its survival and behavior in powdered IFM (Dancer et al., 2009). The temperature range for growth of C. sakazakii is 6°C–45°C (Iversen et al., 2004), with an optimal range of 37°C–43°C. A temperature exceeding the upper growth temperature (heat stress) is encountered by C. sakazakii during the manufacturing process of powdered IFM or its reconstitution in hot water before feeding. When compared with other members of the Enterobacteriaceae family, Cronobacter spp. are reported to be significantly more thermotolerant (Nazarowec-White and Farber, 1997; Breeuwer et al., 2003; Iversen et al., 2004; Edelson-Mammel et al., 2005; Asakura et al., 2007, Chang et al., 2010).
Water activity is an important intrinsic factor in a food that affects bacterial survival and growth. Although dry, powdered IFM has a low water activity (aw 0.2) (Breeuwer et al., 2003), C. sakazakii has been repeatedly isolated from powdered formula (van Acker et al., 2001; Weir, 2002). Compared to other members of the Enterobacteriaceae family, C. sakazakii are reported to be more adapted to dry stress (Caubilla-Barron et al., 2007). Gurtler and Beuchat (2007) studied the survival of C. sakazakii in powdered IFM, and reported that with an inoculum of 2 CFU/g, the pathogen could survive for up to 12 months over a 0.3–0.83 aw range. This finding underscores that Cronobacter spp. may survive in low aw foods such as powdered IFM for extended periods.
A high osmotic pressure is encountered by C. sakazakii during spray drying in the powdered formula manufacturing process and during its persistence in the formula. Recently, Riedel and Lehner (2007) used proteomics tools to identify differentially expressed C. sakazakii proteins in response to osmotic stresses induced by desiccation and growth in hyperosmotic media. Results of their study indicated that under conditions of high osmolarity, various metabolic pathways, including amino acid biosynthesis and transport protein production, were shut down along with the downregulation of the motility apparatus.
Acid stress is another hurdle a pathogen has to overcome in the human stomach. Edelson–Mammel et al. (2006) investigated the acid resistance of 12 strains of C. sakazakii and found that 10 strains showed only a minimal reduction in their counts over 5 h at 36°C in tyrptic soy broth (pH of 3.5). It was also observed that acclimatizing the strains in an acidic medium enhanced the stationary-phase acid resistance, but there was no correlation between stationary-phase acid resistance and thermotolerance. Recently, Lambert and Bidlas (2007) used predictive modeling to study the effects of hurdle technology on the growth and inhibition of Cronobacter spp. and found pH to be an independent inhibiting factor.
Bacteria have evolved adaptive networks to face the challenges of changing environments and to survive under conditions of stress. All adaptive responses involve a series of genetic switches that control the metabolic changes taking place. The σs subunit of RNA polymerase (RpoS) is the master regulator of the general stress response in Escherichia coli and other enteric bacteria (Small et al., 1994; Hengge-Aronis, 1996b). The rpoS regulates response to diverse environmental stresses, including starvation, high osmolarity, and high and low temperatures (Hengge-Aronis, 1996a, 1996b). In turn, rpoS controls expression of many other genes involved in the general stress response such as uspB. Besides rpoS, several genes that specifically respond to heat stress include chaperones such as GroES (Arsene et al., 2000). The PhoP/PhoQ two-component system and the iron regulatory protein Fur, in addition to rpoS, are associated with bacterial gene regulation after exposure to acid stress (Bearso et al., 1997; Gottesman et al., 1997). Similarly, several outer membrane porins such as OmpC, OmpR, and OsmY were found to be upregulated upon exposure to osmotic stress and desiccation in C. sakazakii (Riedel and Lehner, 2007). Despite the availability of a wide body of literature concerning the tolerance of C. sakazakii to a variety of environmental stresses, there is a dearth of knowledge on ways to reduce its stress resistance. Identification of food-grade antimicrobials that reduce stress survival of C. sakazakii would improve the microbiological safety of powdered IFM and help control C. sakazakii infections.
Trans-cinnamaldehyde (TC) is the principal component present in cinnamon oil (Adams et al., 2004). It is a food-grade molecule classified as generally recognized as safe by the Food and Drug Administration. Our laboratory previously reported that TC was effective in inactivating C. sakazakii in reconstituted IFM stored at different temperatures (Amalaradjou and Venkitanarayanan, 2009). The objective of this study was to investigate the efficacy of low concentrations of TC for reducing tolerance of C. sakazakii to environmental stresses such as heat, acid, osmolarity and desiccation, in tryptic soy broth (TSB) and reconstituted IFM. In addition, the effect of TC on the expression of several C. sakazakii genes involved in regulating bacterial stress responses was determined.
Materials and Methods
Bacterial strains, and media
Three strains of C. sakazakii (ATCC 51329, CS 415, and CS 4581) were used for the study. C. sakazakii 4581 was obtained from Dr. Wybo Ingrid, Department of Microbiology, Academisch Ziekenhuis Vrije Universiteit Brussel, Brussels, Belgium. C. sakazakii 415 is a meningitis isolate procured from the Centers for Disease Control and Prevention, Atlanta, Georgia. All bacteriological media used in the study were procured from Difco Becton. Each strain of the pathogen was cultured separately in 10 mL of sterile TSB in 30-mL screw-cap tubes at 37°C for 20 h with agitation (150 rpm). After incubation, the cultures were sedimented by centrifugation (8000 g for 10 min), washed twice, and resuspended in 10 mL of sterile phosphate-buffered saline (pH 7.2). The bacterial population in each culture was determined by plating 0.1-mL portions of appropriately diluted culture on duplicate tryptic soy agar plates (TSA) with incubation at 37°C for 24 h.
Trans-cinnamaldehyde
TC was purchased from Sigma Chemical Co. The TC level used in this experiment was the subinhibitory concentration (SIC) of the compound. The SIC is defined as the highest concentration just below the MIC that does not inhibit growth of C. sakazakii.
Determination of SIC of TC
Sterile 24-well polystyrene tissue culture plates (Costar, Corning Incorporated) containing TSB were inoculated with ∼6.0 log CFU C. sakazakii, followed by the addition of 1–10 μL of TC (Sigma-Aldrich) with an increment of 0.5 μL. The plates were incubated at 37°C for 24 h, and bacterial growth was determined by plating on TSA plates. The highest concentration of TC that did not inhibit bacterial growth after 24 h of incubation was selected as its SICs for this study. Duplicate samples were included and the experiment was repeated three times.
Preparation of IFM
A commercially available brand of powdered IFM was purchased from a retail store and reconstituted as per the manufacturer's instructions. Briefly, 25.5 g of the formula was reconstituted in 180 mL of sterile distilled water, 10 mL volumes were dispensed into 30-mL screw-capped polypropylene tubes, and pasteurized at 63°C for 30 min to remove any inherent bacteria. TC was added to the formula to obtain the aforementioned final concentration.
Effect of TC on C. sakazakii resistance to heat stress
The ability of C. sakazakii to survive heat stress was investigated at 50°C, 55°C, and 60°C (Dancer et al., 2009). Each strain of C. sakazakii was cultured in TSB for 24 h at 37°C and then centrifuged at 5000 g for 10 min at 4°C. The bacterial pellet was resuspended in 10 mL of TSB containing 0 or 750 μM TC and grown to stationary phase by incubation at 37°C for 8 h. Sterile tubes containing 10 mL of TSB or IFM were equilibrated at 50°C, 55°C, or 60°C in a temperature-controlled water bath. At time zero, 100 μL of C. sakazakii culture (6.0 log CFU) was added to the preheated TSB/IFM. At different time intervals, sample tubes or aliquots of samples were removed and immediately cooled in an ice water bath. At 60°C, samples were removed at 2, 4, 6, 8, and 10 min. For 55°C, the samples were removed at 20, 40, 60, 80, and 100 min, whereas at 50°C, aliquots were removed at 30, 60, 90, and 120 min. The tubes were fully submerged in the water bath during the course of the experiment. After cooling, the samples were serially diluted (1:10), plated on TSA, and incubated at 37°C for 24 h before enumeration.
Effect of TC on C. sakazakii resistance to acid stress
To investigate the resistance to acid stress, after growth of C. sakazakii to stationary phase in TSB containing TC (0, 750 μM), the cells were washed with phosphate-buffered saline and added to TSB or IFM (pH adjusted to 3.3) (Chang et al., 2010) at a final population of 6.0 log CFU/mL. The tubes were incubated at 37°C for 0, 10, 20, 30, 40, 50, and 60 min. At each sampling time, the surviving C. sakazakii populations were enumerated as before.
Effect of TC on C. sakazakii resistance to osmotic stress
The effect of TC on C. sakazakii's resistance to osmotic stress was determined according to Chang et al. (2010). Briefly, stationary-phase C. sakazakii cells grown in the presence (750 μM) or absence of TC were washed and resuspended in TSB or IFM containing 75% (w/v) sorbitol (aw 0.81) to obtain an initial population of 6.0 log CFU/mL and incubated at 25°C for 36 h. Samples were removed at 0, 6, 12, 18, 24, and 36 h for enumerating C. sakazakii counts.
Effect of TC on C. sakazakii resistance to desiccation
The effect of TC on C. sakazakii's resistance to desiccation was determined according to the method of Dancer et al. (2009). Stationary-phase cultures of C. sakazakii grown in TSB containing the SIC of TC were washed and resuspended in TSB or IFM, and 100 μL of cell suspension (6.0 log CFU) was transferred to 24-well tissue culture plates. The plates were stored without a lid in a 25°C incubator to air-dry for a 3-week period. At specified time intervals, the inoculums dried on the plate were reconstituted in 1 mL of TSB and appropriate dilutions were plated on TSA. Representative colonies on TSA were confirmed as C. sakazakii based on the characteristic colonial morphology observed when streaked on violet red bile glucose agar plates.
RNA isolation and real-time quantitative polymerase chain reaction
The effect of TC on expression of C. sakazakii genes encoding RpoS, GroES, Fur, PhoP, PhoQ, OmpC, OmpR, Mfla, ClpXP, OsmY, and UspB was investigated using real-time quantitative polymerase chain reaction (RT-qPCR; Sobral et al., 2007). After exposure to the different stresses, C. sakazakii cells were harvested by centrifugation (16000 g for 1 min) at 4°C, and the resulting cell pellet was rapidly frozen in liquid nitrogen and stored at −70°C until RNA extraction. Total RNA from control and TC-treated C. sakazakii, before and after exposure to the aforementioned stresses, was extracted using an RNeasy mini kit (Qiagen) according to the manufacturer's instructions. After extraction, the RNA samples were quantified by spectrophotometry, and stored at −20°C. To eliminate carryover DNA in the RNA extract, 1 μg of RNA was treated for 30 min at 37°C with 150 U of DNase (Qiagen). The DNase was inactivated by heating at 65°C for 10 min. Complementary DNA (cDNA) synthesis was achieved using the Superscript II Reverse transcriptase kit (Invitrogen). The cDNA synthesized was used as the template for RT-qPCR amplification of target genes. The PCR products were detected using SYBR Green. Primers and probes for each candidate gene (Table 1) were designed from published GenBank sequences using Primer Express® software and custom synthesized by IDT USA. RT-qPCR was performed using a 7500 Fast Real-Time PCR system (Applied Biosystems). Triplicate assays were performed with RNA samples isolated from at least two independent cultures of C. sakazakii. Triplicate assays using three independent RNAs confirmed that transcript levels of 16s rRNA were not significantly different (p > 0.05) between the control and TC treated cells (data not shown). Therefore, 16s rRNA was used to normalize the data. The level of candidate gene expression between TC-treated and untreated samples was compared to study relative gene expression, and the effect of TC on each tested gene.
This gene encodes 16s rRNA, which was used to normalize RT-qPCR data.
Statistical analysis
Each experiment was performed in triplicate, with fresh cultures grown for each replication. The mean value and standard deviation were calculated from the data obtained from three separate experiments for each of the stress conditions investigated. In each experiment, two samples were taken at the specific time intervals for enumerating viable C. sakazakii. Data were analyzed using the proc mixed subroutine of Statistical Analysis Software (SAS). Statistical significance was set at p ≤ 0.05.
Results and Discussion
Since there was no significant difference in the efficacy of TC to reduce stress resistance in the three C. sakazakii isolates, only the results obtained with the ATCC isolate (51329) are presented here. Additionally, there was no significant difference (p > 0.05) between C. sakazakii's stress responses in TSB and IFM; therefore, only the results obtained in IFM are presented here. The SIC of TC that allowed C. sakazakii growth as in control was 750 μM. The average initial C. sakazakii population in the wells was ∼6.0 log CFU/mL. After incubation at 37°C for 24 h, ∼8.0 log CFU/mL of C. sakazakii was recovered from all wells, irrespective of control or TC treatment (data not shown). This confirmed that the TC concentrations used in the assay (750 μM) was not inhibitory toward the bacteria.
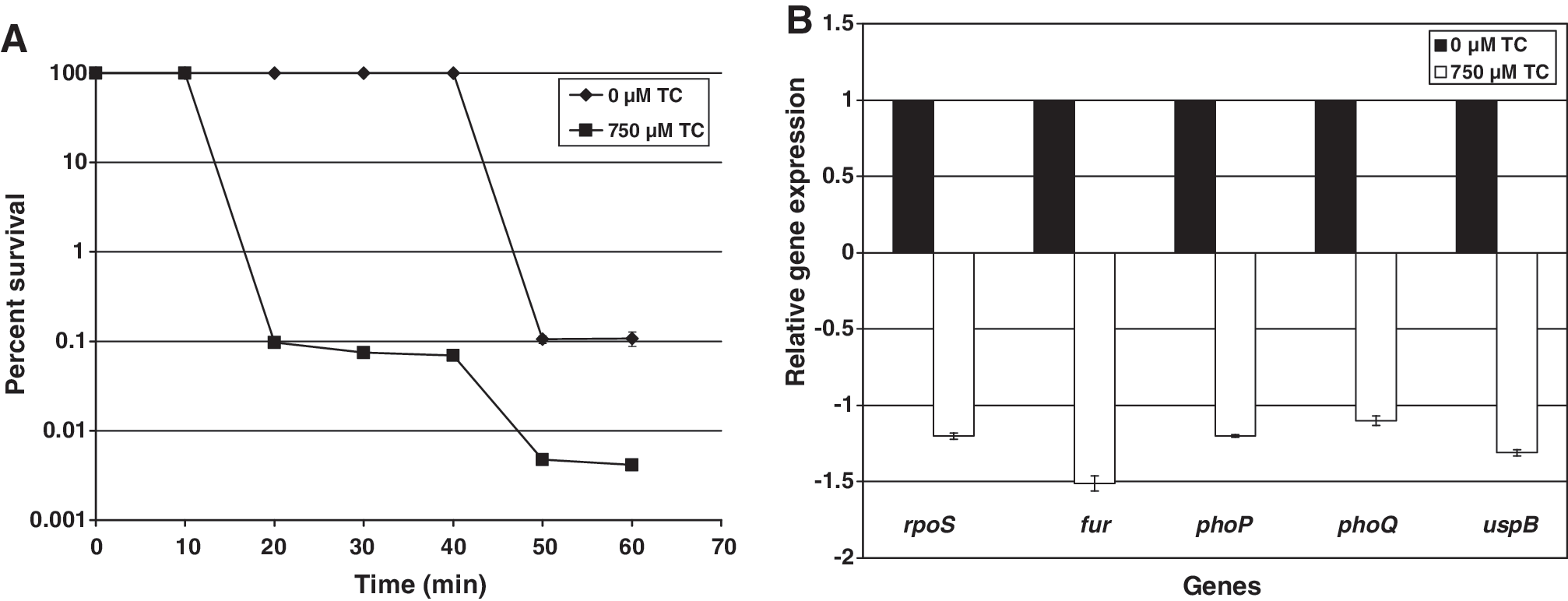
While in the gastrointestinal tract of its host, C. sakazakii must endure the acid pH in the stomach. To determine the effect of TC on reducing acid resistance in C. sakazakii, stationary-phase cells were exposed to an acid challenge of pH 3.3 for a period of 60 min. As depicted in Figure 1A, TC-treated C. sakazakii cells exhibited a diminished ability to survive acid stress compared to control cells (p < 0.05), with more than a 10-fold difference in counts after 60 min of exposure. The RT-qPCR results (Fig. 1B) revealed that TC significantly downregulated expression of phoP/phoQ and fur genes. The role of these genes in regulating the acid tolerance response has been well established in E. coli and Salmonella Typhimurium (Foster et al., 1995; Bearso et al., 1997). In addition, the downregulation of rpoS expression in TC-treated C. sakazakii correlated with the findings of Lee et al. (1995), who showed that rpoS also serves as an acid shock protein in Salmonella Typhimurium.
Heating is commonly employed to control pathogens and spoilage organisms in food so that safety and quality can be preserved. The efficacy of TC in reducing the heat resistance of C. sakazakii at 50°C, 55°C, and 60°C is depicted in Figure 2A–C, respectively. It was observed that at all three temperatures, TC-treated C. sakazakii had a considerably reduced survival compared to the TC-untreated control (p < 0.05) At 60°C, TC-treated C. sakazakii populations had dropped below detectable levels (by plating) at 60 min of exposure (Fig. 2C). Similarly, at 120 and 100 min of heating at 50°C and 55°C, there was ∼1000-fold difference in C. sakazakii counts between TC-treated and control samples (Fig. 2A, B). The efficacy of TC to reduce thermotolerance in C. sakazakii can be explained in part by its gene-modulating effects, as depicted by the RT-qPCR results (Fig. 2D). The downregulation of rpoS expression and upregulation of clpXP could lead to the formation of a less stable RpoS, thereby inhibiting its role as a master regulator of the general stress response (Hengge-Aronis et al., 1996b; Bouche et al., 1998). In addition, there was a downregulation (p < 0.05) of the mfla-1165 gene, which was proposed as a biomarker for thermotolerance in C. sakazakii (Riedel and Lehner, 2007). Another class of proteins expressed after exposure to heat stress in bacteria is the chaperones that aid in proper refolding of misfolded and partially folded proteins (Gottesman et al., 1997). Notably, it was also observed that groES (gene encoding the chaperone GroES) expression was significantly downregulated by TC.
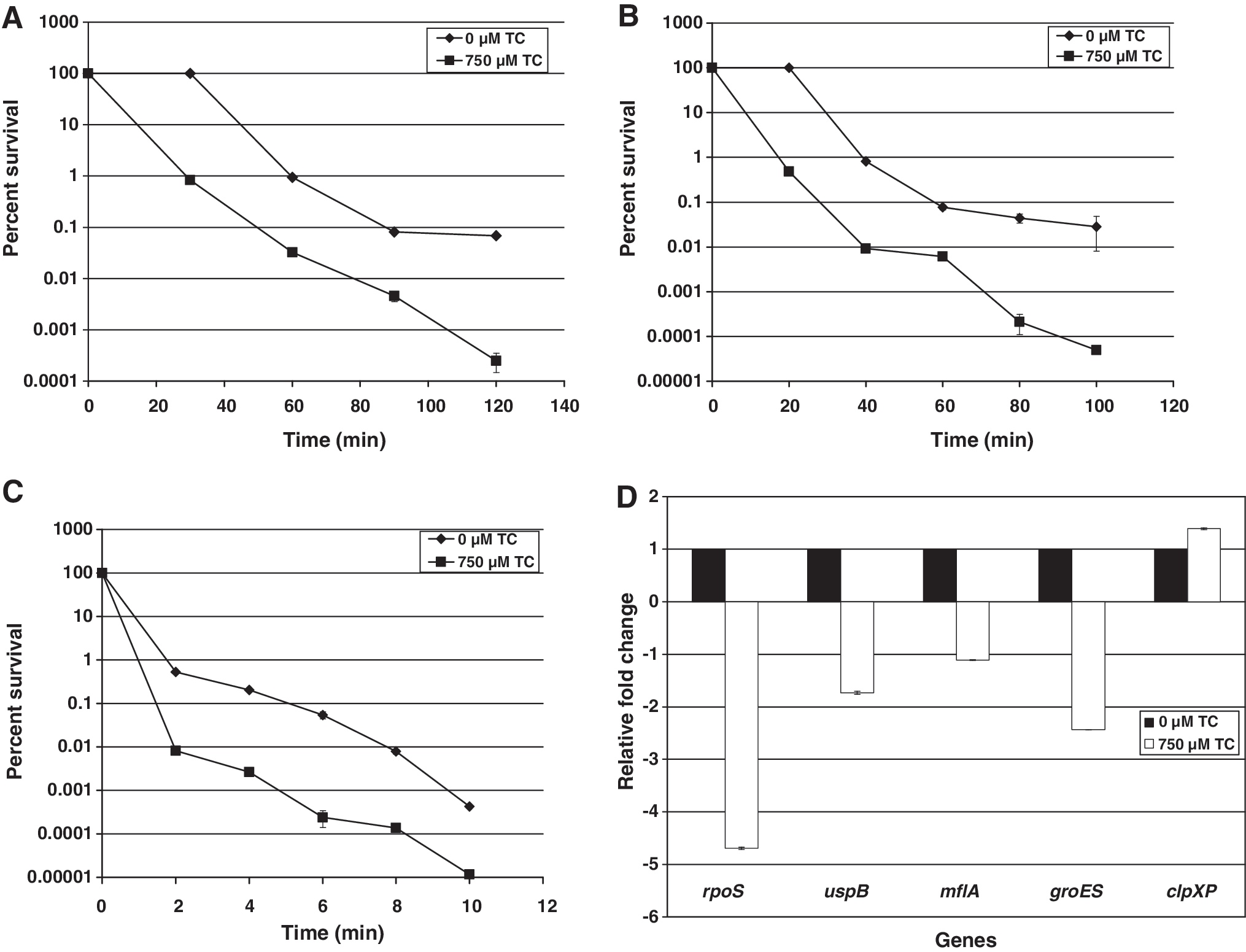
Inhibition of heat stress response in C. sakazakii by TC at 50°C
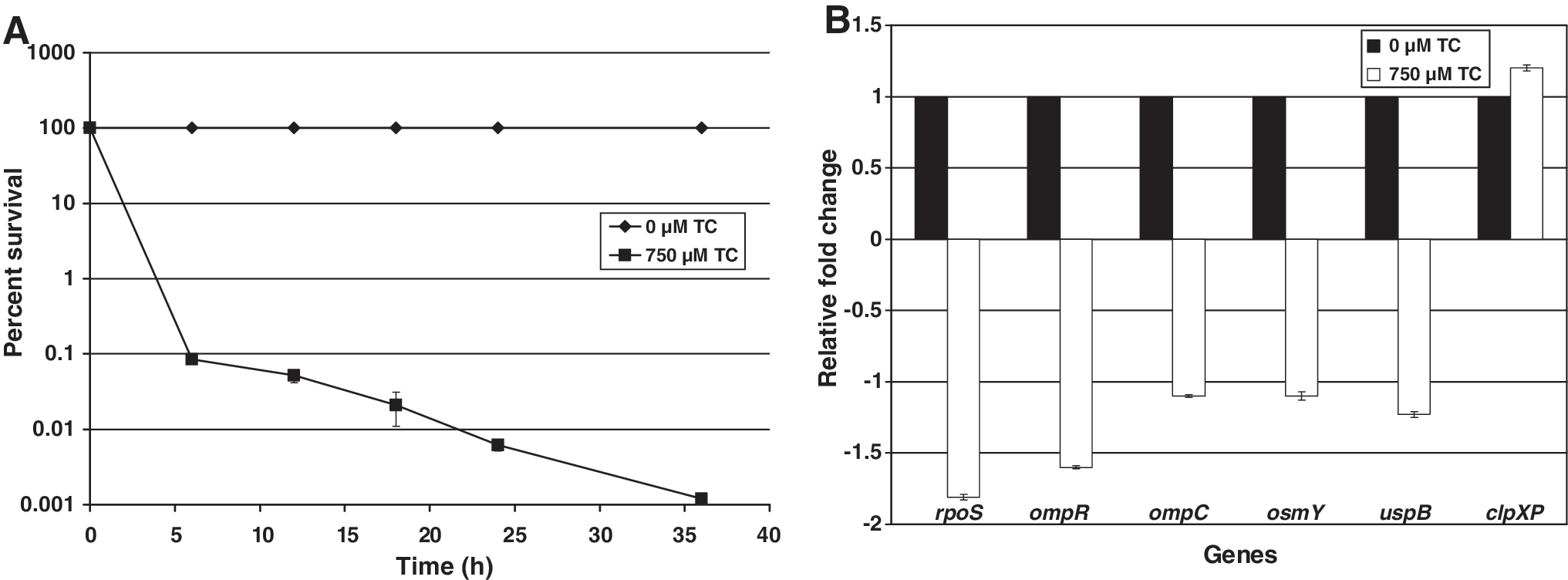
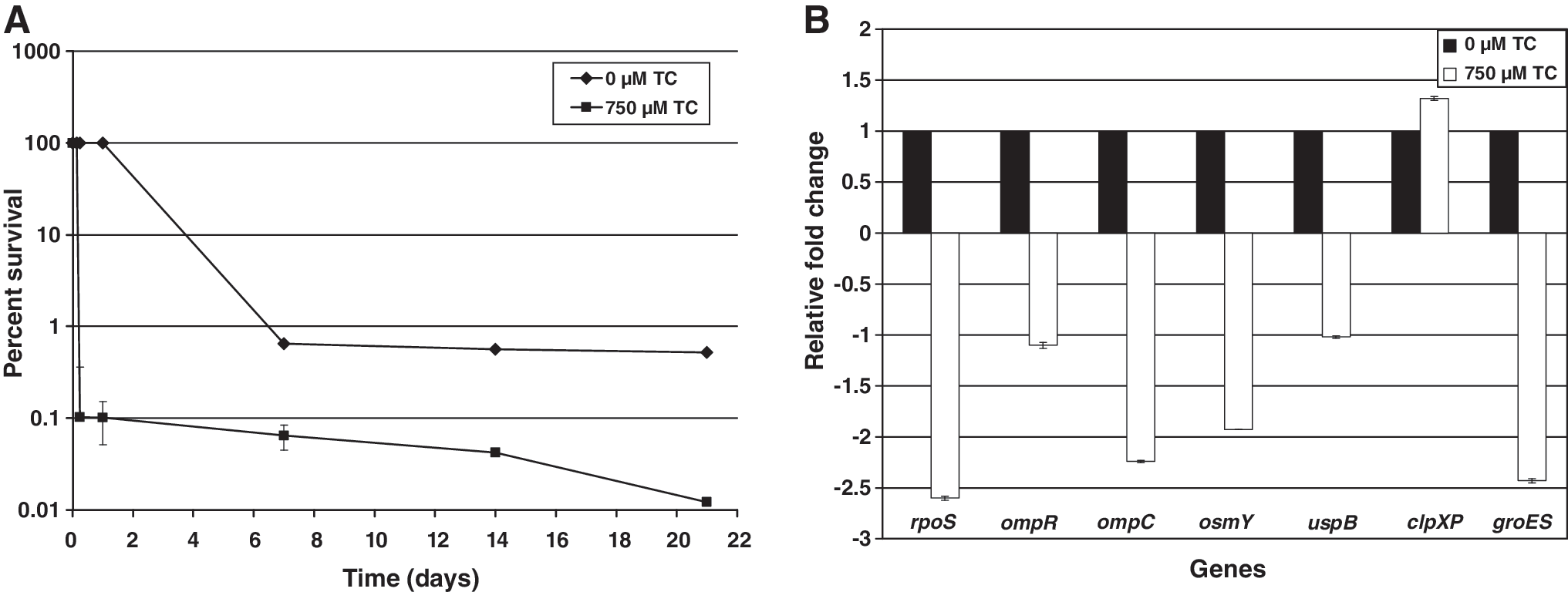
Water activity is an important intrinsic factor of a food that has a significant bearing on microorganisms. Among the various ways to preserve food is the reduction of water activity by increasing its osmotic pressure by addition of high amounts of osmotically active compounds, and by desiccation. Adaptation to osmotic stress is crucial to the growth and survival of C. sakazakii in IFM and processing plants. The efficacy of TC to reduce resistance of C. sakazakii to osmotic stress was investigated in TSB containing 75% sorbitol (aw 0.81). It was observed that TC significantly decreased (p < 0.05) the osmotic stress resistance of C. sakazakii (Fig. 3A). At the end of 36 h, the bacterial counts in TC-treated samples were reduced by greater than 100,000-fold, whereas C. sakazakii populations in controls remained the same, underscoring its ability to tolerate osmotic stress. RT-qPCR studies revealed a downregulation (p < 0.05) of outermembrane porins (OmpC, OmpR, and OsmY; Fig. 3B) that are involved in the transport of osmoprotectants across the cytoplasmic membrane of bacteria under hyperosmotic stress (Pratt et al., 1996; Riedel and Lehner, 2007). Besides reducing resistance to osmotic stress in a liquid medium, TC was also effective in decreasing C. sakazakii's resistance to desiccation. As depicted in Figure 4A, a 10-fold difference in C. sakazakii counts (p < 0.05) between TC-treated and control samples was observed after 7 days of desiccation. The ability of TC to reduce C. sakazakii resistance to desiccation could also be attributed to the decreased gene expression of ompC and ompR (Fig. 4B), which are required for substrate transport across the membrane, and chaperone gene (groES), which is critical for proper folding of misfolded proteins (Osaili and Forsythe, 2009). Gene expression studies using RT-qPCR results revealed that among the various genes tested, rpoS was the only gene that played a role in all the stress conditions imposed on C. sakazakii. This is in agreement with the literature, where rpoS has been reported as a “general stress response regulator” (Lombardo et al., 2004), playing a pivotal function in protecting bacteria under a variety of adverse stress conditions (Hengge-Aronis, 2000; Giuliodori et al., 2007).
In conclusion, TC was effective in reducing stress resistance of C. sakazakii to heat, osmolarity, acidity, and desiccation. Additionally, RT-qPCR results indicated that TC decreased stress resistance in C. sakazakii by downregulating critical genes associated with stress survival. TC could potentially be used as an antimicrobial to reduce its stress tolerance and survivability of C. sakazakii, thereby improving the safety of IFM.
Footnotes
Disclosure Statement
No competing financial interests exist.