
Abstract
Salmonella spp. are important zoonotic pathogens in humans and animals. A longitudinal study was conducted at the Iowa State University's campus (at the Dairy/Animal Science Education and Discovery Facility) to observe change in Enterobacteriaceae (specifically Salmonella) before and after the placement of dairy livestock. To our knowledge, this is the first study that evaluated environmental changes of Gram-negative organisms in a new dairy farm environment. Environmental samples were taken using drag swabs and immediately processed in the laboratory using phenotypic methods (replica plating, the BBL Crystal Identification System for enteric/nonfermenter organisms™, and plating on specialized media/broths). Genotypic methods were also used (BAX PCR™ and pulsed-field gel electrophoresis). Organisms identified as Salmonella were sent to the National Veterinary Services Laboratory (Ames, IA) for confirmatory serotyping. Resistance to antibiotics (ampicillin, nalidixic acid, and tetracycline) was determined by replica plating of Enterobacteriaceae and Salmonella isolates using the guidelines of the National Antimicrobial Resistance Monitoring System and Clinical and Laboratory Standards Institute. The microflora of Enterobacteriaceae changed as cattle were introduced and as time progressed. Additionally, multidrug-resistant isolates began to appear immediately after cattle were introduced (multidrug-resistant isolates were rare prior to introduction of livestock). Variables such as temperature and humidity did not affect the proliferation of bacterial organisms. Seventeen Salmonella isolates were identified as Salmonella London and three isolates as Salmonella Montevideo. Based on pulsed-field gel electrophoresis–generated dendrograms, it is likely that 17 Salmonella London isolates and 3 Salmonella Montevideo isolates are clonal.
Introduction
Bacteria are an important part of the soil microflora because of their abundance, their species diversity, and the multiplicity of their metabolic activities (Ranjard et al., 2000). Traditional microbial culture is considered the established method for the examination of environmental microorganisms (Ingle and Bakland, 1994). Culture methods can detect very small number of cells, and approaches such as formulating the medium to target the growth of specific bacteria (while inhibiting others) can be implemented for more sophisticated and specific measurements (Collinge and Ray, 2006). However, culturing can be time-consuming and take up to 7 days to complete (Singer et al., 2006). Bacteria can also be uncultivable, and viable cells that lose the ability to form colonies may not be detected (Keer and Birch, 2003; Collinge and Ray, 2006). Plate counts assume that during the process of plating, individual colonies are separated and will form discreet colonies; if cells are aggregated, underestimation may occur (Collinge and Ray, 2006). Despite the disadvantages, the ability of microbiologists to cultivate bacteria is still considered extremely important to obtain a thorough understanding and evaluation of microbial communities (Ranjard et al., 2000; Elsas et al., 2007).
During the last 25 years, molecular techniques have increased in prominence, allowing individual bacterial species to be identified and sometimes quantified from environmental samples (Collinge and Ray, 2006; Elsas et al., 2007). Polymerase chain reaction (PCR) technology is used to decrease the time and increase the sensitivity of detection of bacteria in food, environmental, and clinical samples (Singer et al., 2006). PCR is a specific and sensitive technique used to detect nucleic acids; however, the presence of inhibitory compounds may affect PCR and give false-positive results (Denis et al., 2001). Suboptimal reaction conditions (inappropriate primers, improper temperature conditions, incorrect Mg2+ concentration) may also influence the outcome of PCR (Wilson, 1997).
Another DNA-based technology that is used to identify bacterial species and their relatedness is pulsed-field gel electrophoresis (PFGE). PFGE “fingerprint” reflects the structural organization of bacterial chromosome (Prevost et al., 1992) and is the current “gold standard” for highly discriminatory subtyping of most bacterial pathogens (Gerner-Smidt et al., 2006; Stackebrandt, 2006) including Salmonella spp. The disadvantages of PFGE are complex and time-consuming DNA preparation and electrophoresis (Woodford and Johnson, 1998), costly reagents, and the costs of electrophoresis equipment (Mhand et al., 1999). Further, certain organisms such as Clostridium difficile may not be typeable by PFGE because their DNA cannot be extracted intact (Thomas and Weber, 2001).
At the Iowa State University (Ames, IA), a longitudinal study was conducted to observe the change in environmental organisms before and after the placement of dairy livestock (paying particular attention to Enterobacteriaceae and Salmonella spp.). Phenotypic and genotypic methods (such as biochemical requirements of organisms and PFGE profiles, respectively) were utilized to identify and classify organisms. Research described in this article was completed during seven sampling periods from October 20, 2007 to May 28, 2008 (Table 1). Environmental sampling was completed in once monthly intervals during the morning hours (beginning at 9:00 AM Central Standard Time). Sterile drag swabs were used to collect environmental samples; drag swabs have been demonstrated to be effective in sampling poultry (Charlton et al., 2005), swine (Zewde et al., 2009), and cattle (Rahn et al., 1997) facilities. A total of 140 drag swabs were processed in the study; one swab was used for each sampling location for a total of 20 drag swab per sampling date (Fig. 1) and seven sampling dates total (A–G, Table 1).
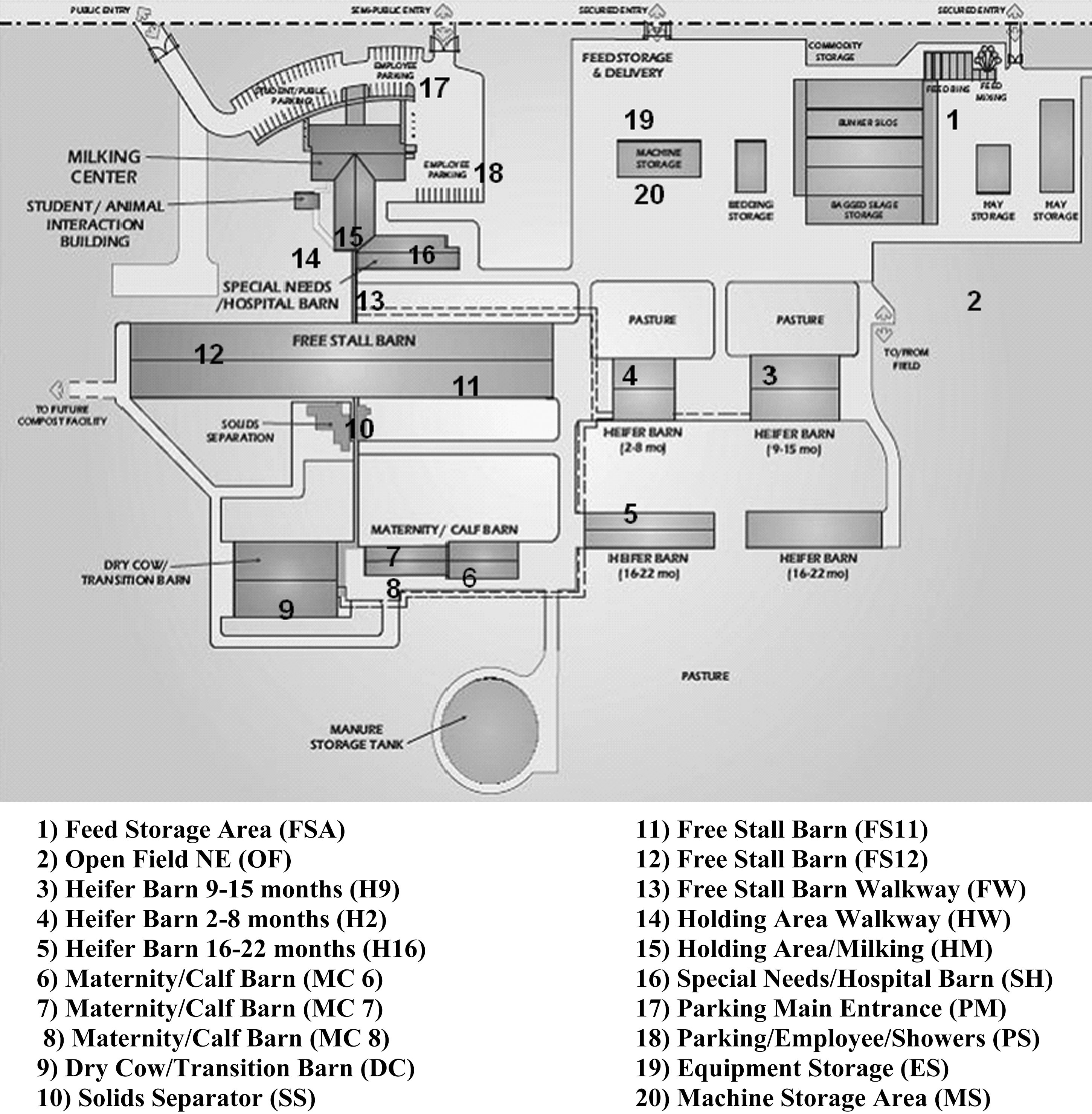
Sampling locations at the Iowa State University Dairy/Animal Science Education and Discovery Facility. (1) Feed storage area (FSA); (2) open field NE (OF); (3) heifer barn, 9–15 months (H9); (4) heifer barn, 2–8 months (H2); (5) heifer barn, 16–22 months (H16); (6) maternity/calf barn (MC 6); (7) maternity/calf barn (MC 7); (8) maternity/calf barn (MC 8); (9) dry cow/transition barn (DC); (10) solids separator (SS); (11) free stall barn (FS11); (12) free stall barn (FS12); (13) free stall barn walkway (FW); (14) holding area walkway (HW); (15) holding area/milking (HM); (16) special needs/hospital barn (SH); (17) parking main entrance (PM); (18) parking/employee/showers (PS); (19) equipment storage (ES); (20) machine storage area (MS).
Materials and Methods
The Iowa State University's Dairy/Animal Science Education and Discovery Facility (henceforth referred to as the “ISU Dairy Farm”) is located on an 887-acre site, 3 miles south of ISU's central campus. The farm houses 450 milking cows plus a similar number of heifers, dry cows, and calves. A complex of buildings were constructed on 27 acres of land, including a free-stall barn, special needs/hospital barn, maternity barn, and a calf research barn for nutritional and husbandry studies. The land the ISU Dairy Farm now occupies was donated to the university by a donor. The farm had been recreational and no livestock had been ever introduced. Dairy cattle were placed on the farm beginning on November 26, 2007 (Table 1). A longitudinal study was conducted to observe the change in environmental microorganisms before and after the placement of dairy livestock (paying particular attention to Enterobacteriaceae and Salmonella spp.). Phenotypic and genotypic methods (such as biochemical requirements of organisms and PFGE profiles, respectively) were utilized to identify and classify organisms. Research described in this article was completed during seven sampling periods (Table 1). Most of the animals were moved from the former ISU Teaching Farm located about 20 miles south of campus (in Ankeny, IA). Additionally, 50 cows were introduced to the new ISU Dairy Farm from a dairy center in Calmar, IA.
During sampling, one sterile 3 cm × 3 cm drag swab moistened with 10 mL skim milk (Solar Biologics, Ogdensburg, NY) was used per sample. Briefly, a drag swab was pulled through the farm environment (Fig. 1) for 60 seconds at normal walking pace (approximately 3 miles per hour). Care was taken to obtain samples in the same locations during subsequent sampling periods. Each sample was placed in a cooler kept at 4°C.
After collection, samples were taken to the laboratory and immediately processed. Each drag swab was aseptically added to a sterile Whirl Pack 24-oz/720-mL filtered homogenizer bag (Nasco, Fork Atkinson, WI) containing 10 mL of buffered peptone water (BPW; Difco, Becton Dickinson Company, Sparks, MD) to add moisture for easier pipetting. The sample was homogenized for 45 seconds at 250 rpm. One milliliter of sample was added to a tube containing 9 mL BPW, creating a dilutent concentration of 10−1. The samples were then serially diluted. A quantity of 0.1 mL dilutent was plated on xylose-lysine-tergitol-4 (XLT-4) agar and incubated for 48 hours at 37°C. Each plate was screened for the presence or absence of black colonies, characteristic of Salmonella spp., and populations of typical Salmonella colonies were enumerated after incubation. The BBL Crystal Identification Systems Enteric/Nonfermerter ID kit™ (Sparks, MD) was used to further identify colonies to the genus and species level.
Resistance to antibiotics was also investigated over time via replica plating. Colonies from XLT-4 media were replica plated (using a replica plating cylinder covered with velvet) onto tryptic soy agar (TSA) (Difco, Becton Dickinson Company). TSA is a general purpose culture medium used for cultivation and isolation of fastidious or non-fastidious microorganisms or for maintenance of stock culture. TSA plate contained ampicillin (48 μg/mL; Sigma, St. Louis, MO), nalidixic acid (48 μg/mL; Sigma), and tetracycline (24 μg/mL; MP Biomedicals, Solon, OH); these quantities were at least 1.5 times the concentrations recommended by the National Antimicrobial Resistance Monitoring System. All antibiotic-containing replica plates were prepared using the guidelines of the Clinical and Laboratory Standards Institute and refrigerated 1–2 weeks before sampling was carried out.
Medications were used to treat conditions such as mastitis and bovine respiratory disease. The medications were prescribed by an ISU faculty veterinarian who disclosed uses and doses given. Sick cattle were treated individually. Additionally, a detailed protocol was strictly followed by the ISU Dairy Farm staff. BAC-STOP udder predip (Esteam Manufacturing, Calgary, Alberta) and Transcend udder predip (IBA, Millbury, MA) were used prior to and after milking. FC-98 Udder Wash (IBA) was also used as a boot sanitizer.
The BAX PCR system™ (Qualicon, Wilmington, DE) was used to screen for Salmonella. The BAX system uses real-time PCR to detect and confirm the presence of target organisms (such as Salmonella) within 24 hours (Raugel, 1999). After a standard enrichment, cells were lysed to release DNA and placed into the BAX system for detection (Cheung et al., 2007). BAX PCR is sensitive, accurate, and rapid (results can be known in as little as 13–24 hours); however, prolonged enrichment is required to detect lower levels of bacterial contamination (Maciorowski et al., 2000). Additionally, false-positive and false-negative results have been reported (Hoorfar et al., 1999; Tomazelli et al., 2008). In this study, a 5 μL aliquot of 10−1 BPW dilutent (taken from specified locations on the dairy farm) was enriched and later added to 200 μL of lysis buffer. After incubation at 37°C for 20 minutes and 95°C for 10 minutes, the lysate was cooled in a cooling block. Fifty microliters of the lysate was combined with a tablet containing PCR reactants (provided in the screening kit) and subjected to real-time PCR in a BAX System Q7 Thermocycler (Applied Biosystems, Wilmington, DE) using the thermocycling program recommended by the manufacturer. Positive and negative controls were used (Salmonella spp., Enterococcus faecalis, and Listeria monocytogenes) during PCR and each 10−1 dilutent was processed for PCR (for a total of 140 PCR reactions completed). Once BAX samples were identified as Salmonella positive, they were later cultured with additional specialized media to identify salmonellae for PFGE and serotyping. Data were analyzed by EpiInfo (version 3.5), a public domain program created by the CDC's PulseNet Program (Atlanta, GA).
Pulsed-field gel electrophoresis
Chromosomal DNA was prepared following the PFGE protocol developed by the CDC PulseNet program (Ribot et al., 2006) with some modifications; agarose plugs were incubated overnight at 37°C using 10 μL of XbaI (5000 U, 10 U/mL; Roche Diagnostics, Mannheim, Germany). PFGE was carried out with a CHEF III Mapper System (Bio-Rad Laboratories, Hercules, CA). The running conditions were as follows: initial switch time, 2.2 seconds; final switch time, 63.8 seconds; 6 V. The induced angle was 120° and run time was 19 hours. Fragments were estimated by comparison with a lambda ladder (Bio-Rad Laboratories) as a molecular marker. The gels were stained with ethidium bromide and visualized under a UV light using a Gel Doc 2000 (Bio-Rad Laboratories). PFGE patterns were analyzed by the BioNumerics software program, version 2.0 (Applied Maths, Kortrijk, Belgium). Dendrograms were generated using the Dice coefficient for similarity and unweighed pair groups method with arithmetic mean clustering.
Results
Environmental microflora results
Salmonella serotypes
Colonies phenotypically identified as Salmonella were sent to the National Veterinary Services Lab (NVSL) for confirmatory serotyping. Twenty were identified as Salmonella spp. (Table 2); of these, 17 were serotyped as Salmonella London and the remaining 3 were identified as Salmonella Montevideo. All salmonellae were found after dairy cattle placement. In this study, the number of Salmonella colonies identified on 10−5 XLT-4 plates (plated directly from the farm environment to XLT-4 agar) were extremely low (Table 3). Of all the XLT-4 plates inoculated with environmental diluents (20 × 10−5 plates per sampling date, seven sampling dates, for a total of 140 plates), only three XLT-4 plates were found depicting colony forming units (CFUs) phenotypically consistent with Salmonella: F10 (SS), F13 (FW), and F16 (SH) (Table 3). All salmonellae were identified in the F group (after dairy cattle were introduced). Black colonies were marked on the XLT-4 plates while replica plating, and antibiotic resistance profiles were determined from those colonies (Table 3). The isolates studied were resistant to ampicillin and tetracycline, but susceptible to nalidixic acid.
Refer to Figure 1 and Table 1: for example, F10 (SS) = sampled on April 23, 2008; location 10, solids separator.
XLT-4, xylose-lysine-tergitol-4; Number XLT (5), number of CFUs growing on XLT-4 10−5 plates; Amp (A), ampicillin; Nal, nalidixic acid; Tet (T), tetracycline; Per, percent; ARP, antibiotic resistance profile.
Most colonies that grew on XLT-4 plates were phenotypically consistent with Enterobacteriaceae such as Escherichia coli (production of yellow colonies). Other Gram-negative bacteria that will grow on XLT-4 plates include Citrobacter, Proteus, and Enterobacter (Corry et al., 2003). When XLT-4 plates were replica plated on TSA agar containing a predetermined amount of antibiotic, the numbers, percentages, and antibiotic resistance profile of each location were recorded (Table 4). There were a total of 2061 colonies that grew on XLT-4 plates during the course of this experiment. Most colonies (667) grew in the E group (sampling date: March 12, 2008); the least (Charlton et al., 2005) were detected during the first sampling period (A group; November 30, 2007). Of those, 667 were resistant to at least one antibiotic (32% of colonies). When studying antibiotics and the numbers of colonies resistant during all sampling periods, there were 791 isolates (38%) resistant to ampicillin, 27 isolates (or 1.3%) resistant to nalidixic acid, and 170 (8%) resistant to tetracycline. Additionally, there was a more detailed review of antibiotic resistance by groups (A–G) and location (1–20) by both the numbers of colonies and resistance (Table 4). BAX PCR results (indicative of Salmonella spp.) are also listed by group and location. Prior to the introduction of cattle, ampicillin resistance was detected in locations A5 and A9 (heifer barn [16–22 months] and the dry cow/transition barn, respectively). Antibiotic resistance was not detected in sampling period B (November 13, 2007). After the animals were introduced (sampling periods C–H), there was an increase in the number of isolates resistant to one antibiotic (ampicillin or tetracycline) as well as multidrug resistance (MDR). Interestingly, resistance was spread throughout the dairy farm and not limited to certain expected locations (such as the free stall barn and special needs/hospital barn).
Refer to Figure 1 and Table 1: for example, A5 = sampled on October 20, 2007; location 5, heifer barn (16–22 months).
XLT-4, xylose-lysine-tergitol-4; Amp (A), ampicillin; Nal, nalidixic acid; Tet (T), tetracycline; NT, nalidixic acid/tetracycline; AN, ampicillin/nalidixic acid; AT, ampicillin/tetracycline; ANT, ampicillin/nalidixic acid/tetracycline; Per, percent; ARP, antibiotic resistance profile.
Resistance to ampicillin only (n = 9) occurs in many of the sampling periods, with the exceptions of B and F (January 27, 2008 and April 23, 2008). Both tetracycline and nalidixic acid/tetracycline resistance were detected after the placement of cattle (Table 4). A combination of ampicillin and nalidixic acid resistance (n = 4) was found in locations D and F; in D3, the heifer barn (H9); in F1, the free storage area (FSA); in F5, the heifer barn [16–22 months] (H16); and in F17, the main parking lot entrance (PM). No ampicillin/tetracycline resistance were detected before the introduction of cattle; however, after introduction, this antibiotic resistance profile (n = 13) was detected in the remaining sampling periods (C–G), all being indoor locations such as the maternity and calf barns (D8 or MC8), free stall barns (E11 or FS11), and special needs/hospital barns (SH). Likewise, the ampicillin/nalidixic/tetracycline profiles were identified after dairy cattle introduction and is spread throughout the farm environment, one (G2) being detected in an open field (OF). Overall, the percentages of nalidixic acid CFUs were small (when compared with ampicillin and tetracycline). Also, it was noted that the numbers of XLT-4 CFUs would change as sampling progressed. Often the numbers would drop, and then percentages would rise and later decrease. When examining tetracycline resistance only, the few colonies that grew exhibited high antibiotic resistance (50% and 100%, respectively). There were similar results in both ampicillin/nalidixic acid-resistant colonies (increase and then decrease) as with the ampicillin/nalidixic acid/tetracycline-resistant colonies. Overall, there were no steady increases (as would be expected) as Enterobacteriaceae became more established.
When examining the BAX PCR results, there is a relationship between the number of detected “Salmonella-positive” isolates (n = 42) and the dairy farm location; most of the BAX-positive results were in locations 12 (FS, the free stall barn; n = 6), locations 11 and 13 (FS11 and FW, the free stall barn and the free stall barn walkway; n = 5), and locations 10, 14, and 15 (SS, the solids separator; HW, the holding area walkway; and HM, the holding area milking; n = 4). The special needs/hospital barn (SH16) reported only 2 BAX Salmonella-positive results. Location 2 (OF, open field north east), location 5 (H16, heifer barn [16–22 months]), and location 9 (DC, dry cow/transition barn) reported negative BAX PCR results during the study.
PFGE results
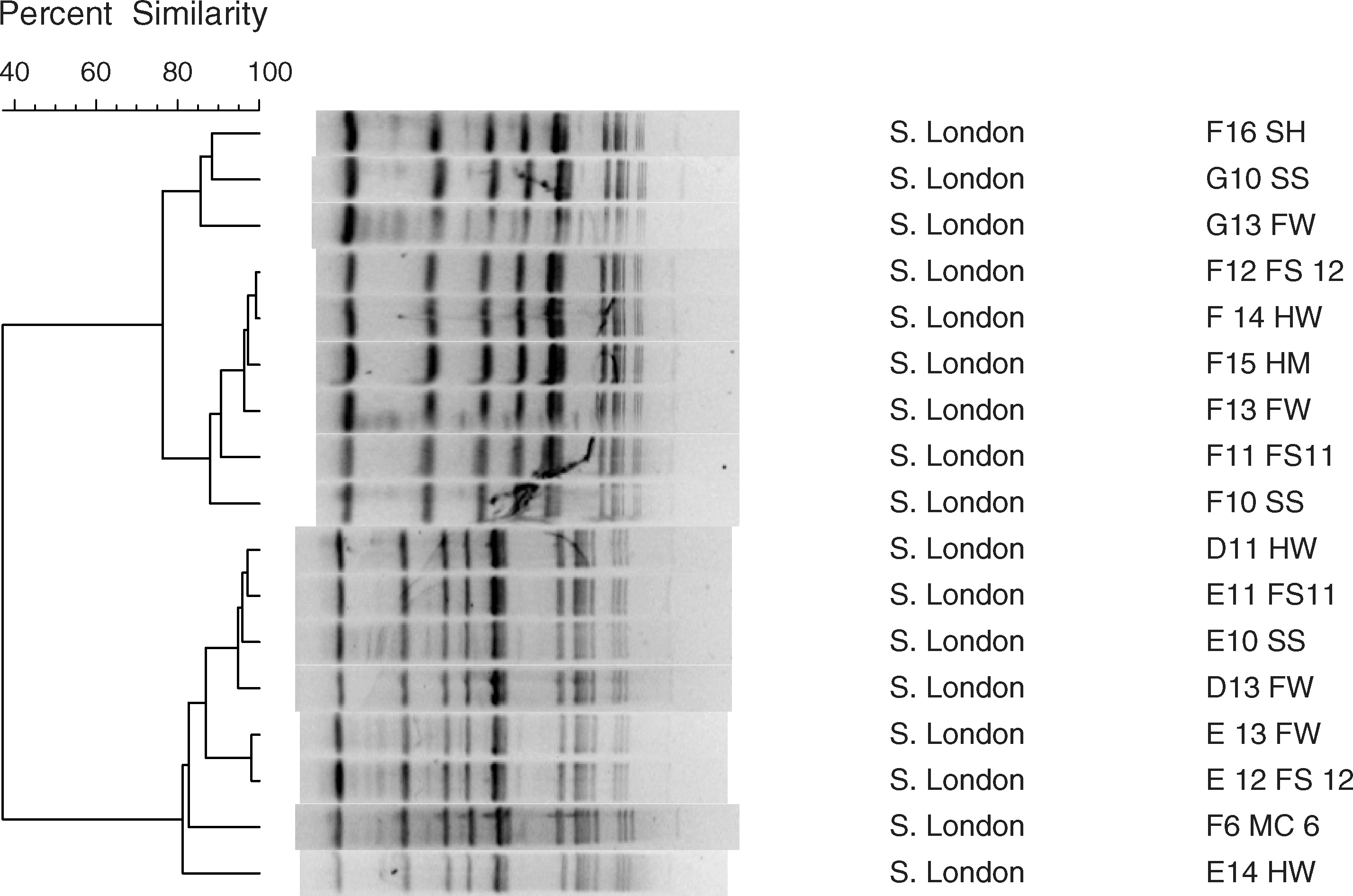
When reviewing the PFGE macrorestriction patterns of Salmonella Montevideo using the criteria of Tenover et al. (1995), there was a one-fragment difference between cluster 1 (F10, F15) and cluster 2 (F14), which is consistent with the gain of a DNA restriction site (data not shown). When examining the PFGE macrorestriction patterns of Salmonella London (also see Fig. 2, lanes 6–9 and 11–13), the patterns appear to be identical, indicating that isolates are indistinguishable from one another (Tenover et al., 1997). When examining Bionumerics-generated dendrograms of the PFGE patterns of Salmonella Montevideo (partial data, Fig. 2, lanes 3 and 4), there were two clusters. The members of cluster 1 included D10 (the solids separator [SS]) and D15 (the holding/milking area [HM]); the only member of cluster 2 was D14 (the holding area walkway [HW]). The PFGE profiles of Salmonella London (Fig. 3) resulted in two distinct clusters: cluster 1 (F16, G10, G13, F12, F14, F15, F13, F11, and F10) and cluster 2 (D11, E11, E10, D13, E13, E12, F6, and E14), both with at least 85% similarity (Fig. 3). All Salmonella isolates were discovered after cattle were introduced on the dairy farm, suggesting that the source of the isolates were likely cattle (Tankson et al., 2006; Magistrali et al., 2008), although there can be other sources of the organism (such as animals and insects attracted to the facility for shelter and food) (Dominguez et al., 2009; Oliver et al., 2009). In PFGE analysis, one or two fragment difference may not indicate a difference in strains. Researchers have suggested that isolates with such minor profile differences are clonally related (defined as isolates recovered from different sources and locations at different times, demonstrating many identical phenotypic and genotypic traits) (Eisenstein, 1990; Thong et al., 1995). When examining the information within both clusters, most of cluster 1 isolates (7/9 or 78%) were collected on April 23, 2008, within the large complex that housed adult animals (free stall barn, special needs/hospital barn, holding areas). The majority of cluster 2 isolates (5/8 or 63%) were collected on March 12, 2008, again within the complex that housed large animals.

Example of a pulsed-field gel electrophoresis gel with Salmonella Montevideo and Salmonella London isolates. Lanes 1 and 15: lambda ladder standard; lanes 3 and 4: D14 and D15 (Salmonella Montevideo); lanes 6–9 and 11–13: Salmonella London D11, D13, E10, E11, E12, E13, and E14, respectively). Also refer to Figure 1 and Table 1: for example, D11 = sampled on February 29, 2008; location 11, free stall barn.
Discussion
Infection with nontyphoidal Salmonella serotypes represents an important public health problem worldwide (Oloya et al., 2009). Salmonella Montevideo has been identified as one of the most common Salmonella isolates found on dairy farms (Wells et al., 2001; USDA, 2005) and has been implicated in outbreaks caused by cheese made from raw milk (Dominguez et al., 2009), bean sprouts (Warriner et al., 2003), and sheep and wild birds (Reilly et al., 1985). In a study of antibiotic susceptibility of Salmonella and U.S. dairy farms by the USDA's Animal and Plant Health Inspection Service, Salmonella Montevideo isolates were sensitive to streptomycin and ampicillin (34% and 9%, respectively), but resistant to tetracycline (USDA, 2005).
Little is known about the rare (but emerging) serotype Salmonella London. Most human cases have been reported in Korea and are typically related to the consumption of food products including powdered milk and other dairy products (Kim et al., 2003; Park et al., 2004). In early reports regarding the antimicrobial susceptibility of Salmonella London, all isolates tested were sensitive to drugs such as chloramphenicol (Sethi et al., 1976). However, Salmonella London is beginning to spread worldwide and has been implicated in products other than dairy-related items (Rowe et al., 2003; Bosilevac et al., 2009). Pet chews, pig meat, meat products, and environmental wastewater have been found to harbor Salmonella London (Wong et al., 2007, 2009; Bosilevac et al., 2009). In addition, there is much variation in the antibiotic resistance of Salmonella London isolates. In a 2006 study of wastewater in Spain, Salmonella London isolates were sensitive to all antibiotics tested (including ampicillin, nalidixic acid, and tetracycline) (Espigares et al., 2006). In a 2009 U.S. study on the prevalence of salmonellae in ground meats, Salmonella London was found to be sensitive to tetracycline but resistant to antibiotics such as ampicillin and nalidixic acid (Bosilevac et al., 2009).
BAX PCR was much more sensitive at detecting salmonellae in environmental samples (n = 42) than initial sampling with XLT-4 (n = 3). In a Swedish study (Eriksson and Aspan, 2007) comparing BAX PCR to other methods of detection (such as modified semisolid Rappaaport-Vassiliadis agar [MSRV] and the selective enzyme-linked immunoabsorbent assay), MSRV performed slightly better than BAX (93% accuracy and 88% sensitive for BAX versus 99% accuracy and 98% sensitivity for MSRV). The selective enzyme-linked immunoabsorbent assay was only 78% accurate and 63% sensitive. Swedish researchers used fecal samples “spiked” with Salmonella, artificially contaminated poultry, swine, and other fecal samples. Other studies have also attested to the high accuracy and sensitivity of the BAX method (Bailey, 1998; Tomazelli et al., 2008; Tice et al., 2009).
The percentage of antibiotic resistance of Enterobacteriaceae (Table 4) did not depend on factors such as temperature and humidity, also referred to “seasonality” in some literature (Haley et al., 2009); however, this study was done over a 9-month period (data not shown). The lack of seasonality has been also supported by studies in the United States (Pradhan et al., 2009) and internationally (Shahada et al., 2008). Conversely, there are studies that depict the seasonality of Salmonella (Edrington et al., 2008; Haley et al., 2009). In the ISU Dairy Farm study, antibiotic-resistant colonies were discovered throughout the sampling periods. However, MDR colonies appeared after cattle were introduced (e.g., ampicillin/nalidixic acid resistance was detected at the dry cow/transition barn [D9]; ampicillin/tetracycline resistance initially appeared at the special needs/hospital barn [C16]; and ampicillin/nalidixic acid/tetracycline resistance was first detected in the maternity/calf barn [C8]). Of note, the percentages of antibiotic resistance varied during sampling periods, as in other studies (Pradhan et al., 2009). There were also intermittent levels of Salmonella (repeated decreases and increases in bacterial levels) during the ISU Dairy Farm study, again similar to results in other studies (Pandya et al., 2009).
Although animals can be infected clinically or subclinically, Salmonella often persists in the subclinical form, making the detection of infected animals difficult (Pandya et al., 2009). Carrier animals may also play an important role in the spread of infections in herds and consequently serve as sources of food contamination and human infection (Zhao et al., 2007). Emergence of MDR Salmonella reduces therapeutic options in cases of infection in humans and animals. MDR resistance is an increasingly important issue worldwide; examples include studies in the United States (Alcaine et al., 2007), China (Cui et al., 2009), and Brazil (Fonseca et al., 2006).
Conclusion
From these experiments (and others cited in this article), there is value in continued monitoring of changes in environmental bacteria in farming environments, particularly changes in antimicrobial susceptibility and resistance. To our knowledge, this is the first study that evaluated the change of Enterobacteriaceae (and specifically Salmonella) before and after the placement of production animals (dairy cattle). This work has demonstrated that organisms of medical importance such as Salmonella have the ability to quickly establish and proliferate on dairy facilities. It would be interesting to continue this longitudinal study to survey ongoing changes in environmental microflora. Additional antibiotics could be used to obtain more extensive antibiotic-resistance profiles. However, more extensive replica plating experiments would likely be prohibitive because of factors such as cost.
Surveillance programs are helpful in documenting (and preventing) the spread of pathogenic bacteria. However, there is concern that passive laboratory-based surveillance is likely to underreport the true incidence of diseases such as salmonellosis in livestock (Zhao et al., 2007; Alexander et al., 2009). Surveillance activities can be combined with additional approaches to decrease the effects of antibiotic resistance, as reported by researchers such as McAllister et al. (2001) and Oliver et al. (2009). They include (1) not using antibiotics to compensate for poor nutrition, poor hygiene, or the lack of immunization; (2) using antibiotics in consultation with a veterinarian; (3) minimizing as much as possible the use of antibiotics considered important for treating human diseases; and (4) using probiotics to compete with potentially pathogenic bacteria (McAllister et al., 2001; Oliver et al., 2009). Research studies such as those examining the association between management type (organic vs. conventional farms) and the presence of resistant bacterial organisms such as Salmonella are now taking place (Alexander et al., 2009). As foodborne pathogens can be a significant threat to human health (Callaway et al., 2003), it is important to study the epidemiology of antibiotic resistance within both animal and human populations.
Footnotes
Acknowledgments
The authors thank Joe Detrick, Kay Christiansen, Bruce Leuschen, D.V.M., and Orhan Sahin, Ph.D., for their assistance. Financial support was provided in part by the Animal Science Food Safety Consortium and the Food Safety Research Laboratories.
Disclosure Statement
No competing financial interests exist.