
Abstract
In September–October 2007, a mixed-serotype outbreak of verocytotoxin-producing Escherichia coli (VTEC) O145:H28 and O26:H11 occurred in the province of Antwerp, Belgium. Five girls aged between 2 and 11 years developed hemolytic uremic syndrome, and seven other coexposed persons with bloody diarrhea were identified. Laboratory confirmation of O145:H28 infection was obtained for three hemolytic uremic syndrome patients, one of whom was coinfected with O26:H11. The epidemiological and laboratory investigations revealed ice cream as the most likely source of the outbreak. The ice cream was produced at a local dairy farm using pasteurized milk. VTEC of both serotypes with indistinguishable pulsed-field gel electrophoresis patterns were isolated from patients, ice cream, and environmental samples. Quantitative analysis of the ice cream indicated concentrations of 2.4 and 0.03 CFU/g for VTEC O145 and O26, respectively. Virulence typing revealed that the repertoire of virulence genes carried by the O145:H28 outbreak strain was comparable to that of O157 VTEC and more exhaustive as compared to the O26:H11 outbreak strain and nonrelated clinical strains belonging to these serotypes. Taken together, these data suggest that O145:H28 played the most important role in this outbreak.
Introduction
As already reported by De Schrijver et al. (2008), a mixed-serotype VTEC outbreak in the province of Antwerp, Belgium, was investigated during September–October 2007. Hypothesis-generating interviews suggested that the outbreak occurred among consumers of ice cream that was prepared and sold at a local traditional dairy farm. Here, we describe the laboratory investigations that allowed us to confirm ice cream as the source of this outbreak and to determine the concentrations of both pathogens in ice cream leftovers. By means of comparative genetic analyses, we tried to gain some insight in the relative contribution of both VTEC serotypes.
Materials and Methods
Laboratory investigation of human samples
Only stools of hospitalized HUS patients were submitted for routine culture. Due to the interval between the first symptoms and recognition of the outbreak, no samples were obtained from patients with uncomplicated diarrhea. VTEC were searched for in stools of five HUS patients using sorbitol-MacConkey (SMAC) and SMAC with cefixime and tellurite. Colony sweeps and isolated colonies were tested for vtx genes using the consensus primer pair MK1 and MK2 (Karch and Meyer, 1989). PCR-positive colonies were biochemically confirmed, and latex agglutination for O145 and O26 (Statens Seruminstitut, Copenhagen, Denmark) was used to confirm the serogroup. PCR-RFLP was applied to identify the flagellar (fliC) types (Machado et al., 2000). Serum samples were analyzed for antibodies against LPS of serogroups O26, O103, O111, O145, and O157 using a slide agglutination assay.
Analysis of environmental samples
Animal and environmental specimens were collected at the dairy farm where the ice cream was produced. Ten bovine fecal samples were collected: two samples from young calves (<3 months), five from young cattle (±1 year), and three from adult cows. For each pen the bedding was sampled using the overshoes method (Cobbaut et al., 2008). Dust and feed samples were also collected in the different animal houses. The milk tank (one sample) and six batches of ice cream made from pasteurized milk produced at the dairy farm were sampled. Investigators obtained ice cream leftovers from one of the cases for VTEC detection.
Non-O157 VTEC were searched for using the isolation method described before (Possé et al., 2008b). Briefly, samples (10 g for feed or dairy samples and 1 g for dust) were diluted in 1:10 tryptic soy broth (TSB) supplemented with 8 mg/L novobiocin and 16 mg/L vancomycin. After 6 hours of pre-enrichment at 37°C, 2 mg/L rifampicin, 1.5 g/L bile salts, and 1.0 mg/L potassium tellurite were added to the medium that was further incubated for 18 hours at 42°C. Fecal samples (25 g portions 1:10 diluted using TSB) were directly enriched during 24 hours at 42°C in modified TSB with 8 mg/L novobiocin, 16 mg/L vancomycin, 2 mg/L rifampicin, 1.5 g/L bile salts, and 1.0 mg/L potassium tellurite. After incubation for 24 hours, 100 μL of each enrichment broth was plated onto the recently developed differential agar media for detection of O26, O103, O111, and O145 serogroups (Possé et al., 2008a). In addition, immunomagnetic separation (IMS) was applied after 6 and 24 hours of incubation of the broths. Serogroup-specific beads targeting O26 and O145 antigens (Invitrogen Dynal AS, Oslo, Norway) were applied according to the manufacturers' instructions. Afterward, 100 μL of the IMS suspension was plated onto the differential agar media. These plates were incubated for 24 hours at 37°C. Suspected colonies were transferred to serogroup-specific confirmation media: isolates with a suspected morphology on both differential and conformational media were confirmed using PCR detecting the presence of vtx- and O-antigen-encoding genes (Possé et al., 2007) and slide agglutination with anti-O145 and -O26 antibodies.
Quantification of ice cream contamination
To estimate the level of contamination of the ice cream leftovers, two quantification methods were used. On one hand, quantitative data were obtained by the three tube most-probable-number method (10, 1.0, and 0.1 g) using the isolation procedure described above. On the other hand, quantitative data were obtained by plating 1 g of ice cream onto Rapid E. coli II medium (BioRad, Hercules, CA). Briefly, 10 g of ice cream was diluted using 90 mL TSB. From this dilution, 10 portions of 1 mL each were plated onto Rapid E. coli II medium and incubated for 24 hours at 42°C. All present E. coli–like colonies on these agar plates were further tested by the same methods as the other suspected colonies obtained during the isolation procedures.
Pulsed-field gel electrophoresis
Pulsed-field gel electrophoresis (PFGE) according to the Pulse-Net U.S.A. protocol for E. coli (Centers for Disease Control and Prevention, available at
Virulence profiling of VTEC strains
The virulence profiles of the outbreak strains were investigated using PCR. For comparison purposes, 10 O145 and 12 O26 fecal VTEC, recovered from nonrelated patients at the Belgian Reference Laboratory for VTEC/EHEC between 1991 and 2008, were randomly selected from our cryocollection. The established virulence markers vtx1, vtx2, eaeA, and ehxA were searched for according to the method described by Paton et al. (1998). Additional PCRs targeting plasmid genes for Shiga toxin–producing E. coli auto-agglutinating adhesin (saa), a subunit of subtilase cytotoxin (subA), extracellular serine protease (espP), catalase-peroxidase (katP), and type II transporter system (etpD) were performed (Brunder et al., 1999; Schmidt et al., 1999; Paton and Paton, 2002; Paton et al., 2004). All strains were screened for the presence of pathogenicity island O Island 122 (OI-122) using primers described by Karmali et al. (2003) and Wickham et al. (2006). OI-122 contains 26 open-reading frames (ORFs) that are organized in 3 modules (Wickham et al., 2006). Strains with positive PCR results for 9 selected ORFs were defined as strains carrying a complete OI-122 (COI-122), an incomplete OI-122 was appointed to strains with a negative PCR result for at least 1 OI-122 ORF, and strains with no positive OI-122 PCR result were labeled as “OI-122 absent.” VTEC O157:H- E32511 was used as a positive control in all PCRs, except for saa and subA for which a clinical VTEC O113:H21 isolate (strain EH1516) was used. PCR-grade water was used as negative control.
Results
Laboratory analysis of human, cattle, and dairy samples
Evidence of VTEC infection was detected in three out of five HUS cases (Table 1). vtx2-positive VTEC O145:H28 and vtx1-positive O26:H11 were isolated from feces and urine of a 2-year-old girl, whereas only O145:H28 was isolated from the feces of an 8-year-old girl. A positive serology test for anti-O145 antibodies was obtained in a third case (an 11-year-old girl). VTEC of serotypes O145:H28 and O26:H11 carrying vtx2 and vtx1, respectively, were isolated from cattle and dairy samples (Table 2). O26 VTEC was isolated after IMS enrichment of bedding samples collected in the stable housing young calves (<3 months). O145 VTEC were recovered from hay and feces of one of the young calves with and without the use of IMS, but from dust only after IMS enrichment. O26 and O145 VTEC were also isolated from ice cream leftovers. No VTEC were isolated from the tank milk and ice cream collected at the farm and from the stable housing adult cattle. PFGE analysis confirmed that VTEC O145:H28 and O26:H11 isolated from patients, ice cream, and environmental samples, respectively, showed indistinguishable patterns (Fig. 1).
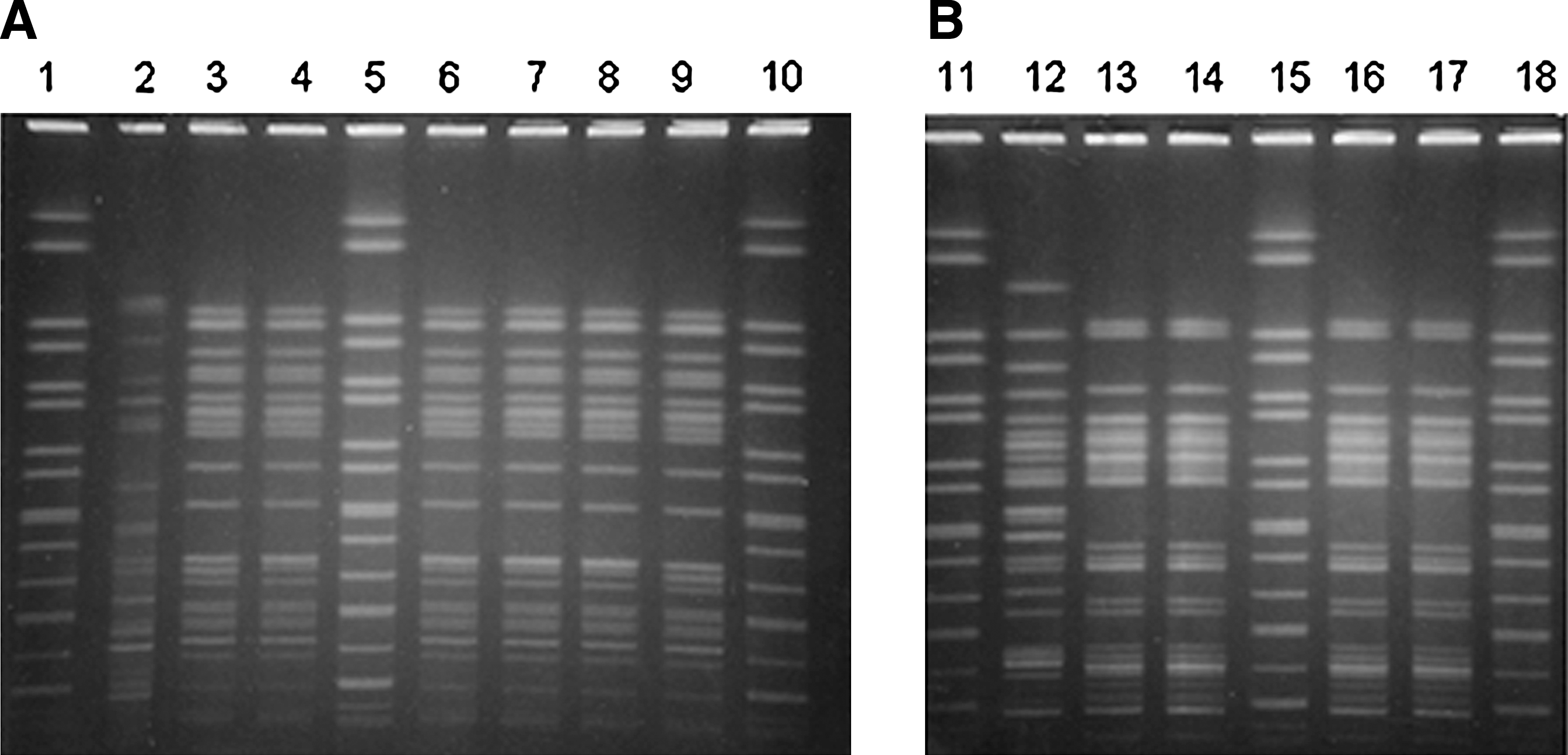
PFGE analysis of VTEC O145:H28 and O26:H11 isolates associated with the HUS outbreak in September 2007. Reprinted from De Schrijver K et al. (2008) with permission.
HUS, hemolytic uremic syndrome; VTEC, verocytotoxin-producing Escherichia coli.
PCR assays aimed at detecting O26, O145, vtx1, and vtx2 genes in samples with suspected colonies.
IMS, immunomagnetic separation; NA, not applicable; vtx, verocytotoxin.
The standard most-probable-number analysis on ice cream leftovers showed a count of 2.4 and 0.03 CFU/g for VTEC O145 and O26, respectively. Quantitative data obtained after direct plating of 1 g of leftover ice cream on Rapid E. coli II medium resulted in 20 E. coli isolates. Out of these 20 isolates, 2 were confirmed as VTEC O145 using PCR, resulting in a concentration of 2.0 CFU/g. Plating onto Rapid E. coli II medium did not allow the isolation of VTEC O26 because of the detection limit (1 CFU/g) of this method.
Virulence profiling of outbreak and nonrelated strains
All O145 and O26 outbreak and nonrelated sporadic strains showed H28 and H11 patterns after fliC PCR-RFLP, respectively. The O145:H28 (EH1533) and O26:H11 (EH1534) outbreak strains were positive for vtx2 and vtx1, respectively (see Supplementary Table S1; Supplementary Data are available online at
The O145:H28 outbreak strain carried a COI-122 with 9 ORFs present (see Supplementary Table S1). In contrast, no PCR amplification was obtained for the 3 ORFs belonging to Module 1 and ORF Z4323 of Module 2 in the O26:H11 outbreak strain resulting in an incomplete OI-122, and possibly a less pathogenic VTEC. Among the sporadic isolates, one O26:H11 (strain EH296) possessed a COI-122. The majority (19/22) of strains contained an incomplete OI-122, all were PCR negative for Module 1 and ORF Z4323, and OI-122 was completely absent in one O145:H-.
Discussion
We have described the laboratory investigations of a mixed-serotype outbreak of VTEC infections among consumers of ice cream produced at a farm in Belgium. To our knowledge, this is the first mixed-serotype outbreak involving VTEC serotypes O145:H28 and O26:H11. Simultaneous infections of O157 and non-O157 VTEC have been described previously (Rivas et al., 1993; Ludwig et al., 1996) and a case of HUS following coinfection of O177:H- and O55:H7 was recently reported (Gilmour et al., 2007). The occurrence of mixed-serotype VTEC infections has long been recognized (Goldwater and Bettelheim, 1996; Bettelheim, 2007). It was suggested that a number of outbreaks ascribed to VTEC O157 may well have been due to other VTEC that were not sought for. In this outbreak, exhaustive testing of single colonies with PCR targeting vtx genes led to isolation of both serotypes from human samples. To detect both VTEC O157 and non-O157, culture-based methods using selective and differential media and simultaneous detection of toxins or toxin genes in stools were recommended by the Centers for Disease Control and Prevention (Gould et al., 2009). PCR is, however, not available in most microbiological laboratories where often only culture-based identification techniques not capable of differentiating non-O157 VTEC are used. This underlines the importance for sentinel laboratories to refer not only isolates to reference laboratories for further investigation, but also positive stools.
This report illustrates the applicability and sensitivity of a new isolation method for the detection of non-O157 VTEC in naturally contaminated dairy products, cattle feces, and environmental samples (Possé et al., 2008a,b). Sensitive methods are required for screening of foods and feces, as contamination can occur at very low concentration. A low infectious dose for VTEC O157 infection, in the order of 100 CFU or less, has been estimated from previous outbreak investigations (Griffin, 1995; Teunis et al., 2004). No data are available concerning the infectious dose of non-O157 VTEC. We showed that ice cream contaminated with a low concentration of O145:H28 (2.4 CFU/g) and O26:H11 (0.03 CFU/g) was able to cause an HUS outbreak. Although no accurate dietary history was obtained from the HUS cases, an average consumption of 200 g ice cream, corresponding to approximately 400 CFU of ingested VTEC O145:H28, was estimated.
Multiple reasons might explain why O145:H28 infection was confirmed in 3 of the outbreak cases, whereas O26:H11 only in one. First, we showed that the concentration of the VTEC O145 strain in ice cream leftovers was 100-fold higher than that of the O26 strain. Second, in contrast to the O26 outbreak strain, the O145 outbreak strain contained a virulence profile associated with an increased risk of HUS development (Karmali et al., 2003; Brooks et al., 2005; Tarr et al., 2005; Wickham et al., 2006). Although both VTEC serotypes have been associated with HUS and strains producing only vtx1 and lacking COI-122 have been recovered from HUS patients, it has been suggested that vtx2 and COI-122, possibly in a synergistic manner, might enhance the virulence potential of VTEC strains.
In conclusion, we have demonstrated that VTEC of non-O157 serotypes, in this case O145:H28 and O26:H11, may be responsible for foodborne outbreaks of HUS. The fact that ice cream produced from pasteurized milk was the vehicle of transmission highlights the concerns about food safety on dairy farms that produce and commercialize their own dairy products on site. Moreover, the low infectious dose at which these strains were able to cause disease and the O145:H28 virulence profile, which is comparable to O157 VTEC, indicates the potential of non-O157 VTEC as human pathogens. Virulence profiling and quantification of non-O157 VTEC could become useful tools for public health services to assess the pathogenicity of these foodborne bacteria and to control their disease burden on society.
Footnotes
Acknowledgments
Strains EH1533 and EH1534 have been deposited in both ATCC (BAA-1652 and BAA-1653) and BEI Resources (NR-10147 and NR-10148). This research was supported by the Institute for the encouragement of Scientific Research and Innovation of Brussels grant PRFB 2007-29. Part of this work was presented at the Pathogenic Escherichia coli Network Conference, “Epidemiology and Transmission of VTEC and other Pathogenic Escherichia coli,” Stockholm, 25–26 September 2008.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.