
Abstract
The foodborne pathogen Listeria monocytogenes is able to form biofilms in food processing environments. Previously, we have reported that an lm.G_1771 gene (encoding a putative ABC-transporter permease) was involved in negative regulation of L. monocytogenes biofilm formation using LM-49, a biofilm-enhanced mutant isolated on Tn917 mutagenesis (AEM 2008 p.7675–7683). Here, the possible action of this ABC-transporter permease in L. monocytogenes biofilm formation was characterized by phenotypic, proteomic, and genomic analyses using an lm.G_1771 gene deletant (Δ1771). The Δ1771 mutant exhibited the same enhanced ability for biofilm formation as the LM-49 strain using a crystal violet staining assay. DNA microarrays and two-dimensional gel electrophoresis revealed 49 and 11 differentially expressed (twofold or more) genes or proteins in Δ1771, respectively. The transcriptomics study indicated that lm.G_1771 could play a vital role in regulating candidate genes involved in biofilm formation such as genes encoding cell surface proteins (Dlt), cell surface anchor proteins (SrtA), and transcriptional regulators (GntR) contributing to negative reglution of biofilm formation by L. monocytogenes. The mutant Δ1771 was more sensitive to Triton X-100 and less resistant to cationic antibiotics, which might be explained by the down-regulation of dlt operon in this deletant and the fact that dlt involves the incorporation of D-alanine residues into lipoteichoic acids, resulting in a positive net charge on the teichoic acids. Therefore, lm.G_1771 is considered to be involved in negative regulation of biofilm formation, and the results from this work provide a possible molecular mechanism of biofilm formation regulated by lm.G_1771 in L. monocytogenes.
Introduction
Bacterial biofilm formation is a complex process involving a number of biosynthetic pathways, and the process is subject to regulation by different signal transduction pathways. In L. monocytogenes, several genes or gene products are implicated in biofilm formation. First, motility proteins such as flagellins are important for bacterial adhesion onto a surface, the initial step in biofilm formation. It has been reported that flagellated L. monocytogenes cells readily colonized on glass or stainless steel, whereas adhesion by a nonflagellated mutant was markedly reduced (Vatanyoopaisarn et al., 2000). Second, two bacterial signal transduction pathways were shown to regulate L. monocytogenes biofilm formation (Ren et al., 2004). One involves the autoinducer 2 (AI-2) universal quorum sensing signalling molecule that is synthesized and transported by well-conserved pathways in bacteria. A key enzyme in this pathway is the AI-2 synthase, which is encoded by the luxS gene identified in the L. monocytogenes EGD-e genome (Ren et al., 2004). A luxS mutant exhibited a biofilm-positive phenotype in L. moncytogenes (Sela et al., 2006), which has also been found in several other bacterial species (Schauder and Bassler, 2001). The other signal transduction pathway is the accessory gene regulator (agr) system, which controls the expression of several virulence factors in Staphylococcus aureus (Shirtliff et al., 2002). This system also regulates biofilm formation in L. monocytogenes, which was demonstrated with mutants with either agrA or agrD deleted from the agrBDCA operon showing markedly reduced adhesion to an abiotic surface (Rieu et al., 2007). Third, a recent study by Harmsen et al. (2010) indicated that extracellular DNA was an important component for L. monocytogenes initial attachment and early biofilm formation through interactions with peptidoglycan (specifically N-acetyl glucosamine).
Recently, we characterized a Tn917 insertion mutant LM-49 and identified that the mutant gene lm.G_1771 (encoding an ABC-transporter permease) was involved in negative regulation of biofilm formation by L. mococytogenes (Zhu et al., 2008). The purpose of this study is to further determine the role of the lm.G_1771 gene in biofilm formation using an lm.G_1771 deletion mutant Δ1771 and its parent L. monocytogenes 4b G by genomic, proteomic, and phenotypic analyses. This research may provide insight into the possible mechanisms of biofilm formation by L. monocytogenes.
Materials and Methods
Bacterial strains, media, plasmids, and growth conditions
The L. monocytogenes 4b G strain was obtained from the Center for Disease Control of Hubei Province, China. The biofilm-positive mutant LM-49 was generated by Tn917 mutagenesis (Chen and Shi, 2005), and the lm.G_1771 deletion mutant (Δ1771) was constructed according to the procedure described by Rieu et al. (2007). L. monocytogenes strains were grown in brain heart infusion broth (BD Company) or trypticase soy broth (Merck) at 37°C without shaking.
Preparation of protein samples and proteomic analyses
Total protein preparation for L. monocytogenes and two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) were performed as described by Tremoulet et al. (2002). Protein samples were prepared from three independent cultures for each strain, and each sample was analyzed twice.
The scanned images for stained 2D gels were analyzed with the ImageMaster 2D Platinum software (release 5.0; Amersham Biosciences). Only the spots that exhibited significant differences between the mutant Δ1771 and the wild-type strain in at least n-1 gels (n representing the number of gels run for each condition) were selected for protein identification, and the significance was analyzed using the Student's t-test analysis (95% confidence interval).
Differentially expressed proteins were identified using a matrix-assisted lazer desorption ionization tandem time of flight (MALDI TOF/TOF) instrument (4800 plus analyzer; Applied Biosystems) as previously described (Lametsch et al., 2002). Total spectra were generated and used to search the Swiss-Prot database for bacterial species by using MASCOT (Matrix Science), which revealed the identities of the differentially expressed proteins.
RNA isolation and sample preparation for NimbleGen GeneChip analysis
Three independent overnight cultures of L. monocytogenes 4b G strain and Δ1771 were grown in brain heart infusion medium and were harvested for RNA isolation. The preparation of total RNA was performed as previously described (Zhu et al., 2008). Double-stranded cDNA synthesis, labeling, hybridization, and washing were carried out according to the NimbleGen Array gene expression analysis protocol (Roche, Mannheim, Germany). The cDNA labeled either with Cy3 or Cy5 was hybridized to a NimbleGen 4 × 72K 45-60-mer microarray slide (
Real-time quantitative polymerase chain reacton
The preparation of total RNA was the same as for microarray analysis. One hundred ng of total RNA were added to a 25-μL polymerase chain reacton (PCR) reaction mixture employed for quantitative PCR according to the instructions provided with the One Step SYBR PrimeScript RT-PCR Kit (TaKaRa). Bio-Rad iCycler (Bio-Rad Laboratories) was used to detect fluorescence with the following protocol for the PCR: one cycle at 42°C for 5 min (reverse-transcript step) and 40 cycles at 95°C for 5 sec and 60°C for 31 sec. The PCR primers used for quantitative PCR are listed in Table 1. 16S rRNA gene fragments were amplified as an internal control, and three biological replicates were performed. Relative gene expression was calculated using the comparative critical threshold cycle (ΔΔCT ) method with PCR efficiency (Livak and Schmittgen, 2001).
Primers designed using Beacon Designer 7.0 software.
Results
Biofilm formation by the lm.G_1771 deletion mutant
To eliminate any possibility that the transposon vector (on LM-49) might contribute to the observed results and to generate a more suitable mutant for further study, an lm.G_1771 in-frame deletion mutant Δ1771was screened. The presence of lm.G_1771 mRNA was determined by the reverse transcription–PCR. The result showed that the lm.G_1771 mRNA was absent in Δ1771 but present in the wild-type strain (data not shown). Physiological characterization of this mutant indicated that it maintained the same cell and colony morphologies as well as growth rate as the wild-type strain (data not shown). However, observations on Δ1771 biofilm formation using the microplate crystal violet staining assay revealed that the deletion mutant exhibited an enhanced level of biofilm formation relative to the wild-type strain, which was consistent with the result obtained for the transposon mutant LM-49 (Fig. 1). This reinforced the conclusion that lm.G_1771 was involved in negative regulation of biofilm formation in L. monocytogenens, and the presence of Tn917 did not influence regulation.
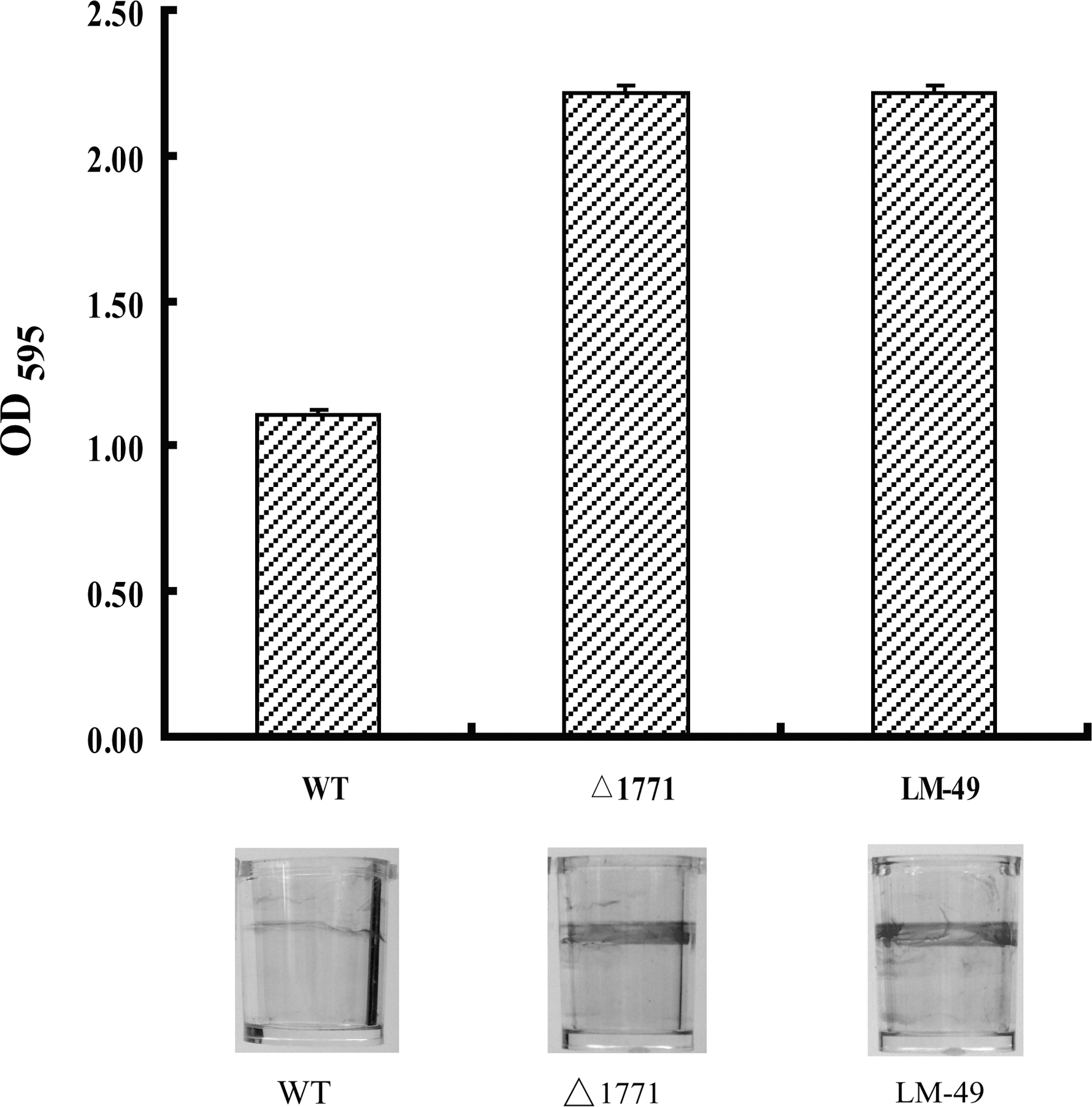
Biofilm quantification using crystal violet staining. Pictures of biofilms formed in microplates were shown below each column. WT: wild-type strain; Δ1771: lm.G_1771 deletion mutant; LM-49: Tn917 insertion mutant.
Identification of proteins differentially expressed in Δ1771 using 2D-PAGE and MALDI-TOF/TOF mass spectrometry
To gain insight into the mechanisms by which lm.G_1771 regulates biofilm formation, we examined differential protein expression in the deletion mutant and the wild-type strain. Approximately, 800 protein spots were obtained by 2D-PAGE from each protein gel using the ImageMaster 2D Platinum software (Fig. 2). Comparing the protein pattern of the Δ1771 mutant with that generated from the wild-type strain resulted in the identification of 15 differentially expressed proteins in the mutant, including 11 up-regulated proteins and 4 down-regulated ones (two- to eightfolds). Eleven of the protein spots were identified by MALDI TOF/TOF mass spectrometry performed after an in-gel trypsin digestion (Table 2). Unexpectedly, the fourth down-regulated protein spot in Δ1771 (i.e., the Lm.G_1771 permease) was not found in the wild-type by 2D-PAGE. One possible explanation was that the predicted isoelectric point (pI) of this Lm.G_1771 protein, which was 9.09, was beyond the pH range (pH 4–7) of the immobilized pH gradient (IPG) strips used.
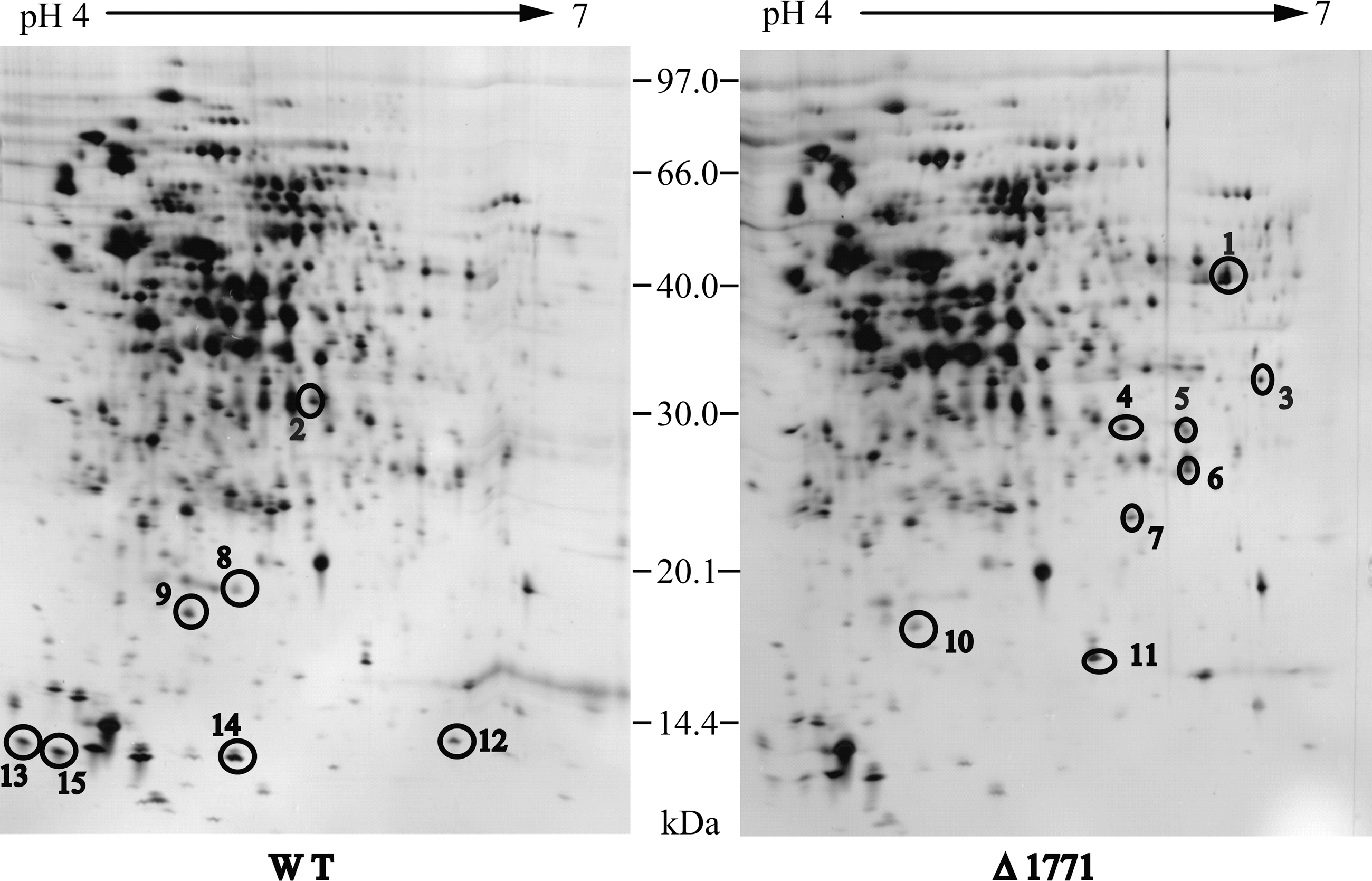
Images of the silver-stained two-dimensional protein gels of Listeria monocytogenes. Circles and numbers indicate spots increased more than twofold between the wild-type L. monocytogenes and the Δ1771 mutant. The approximate positions of the sodium dodecyl sulfate-polyacrylamide gel electrophoresis molecular mass (MW) standards are presented between the gel images. The proteins that were identified are described in Table 3.
Spot number according to the position on the two-dimensional polyacrylamide gel electrophoresis (Fig. 3).
Protein entries in National Center for Biotechnology Information.
Protein Score based on MOWSE Score (Mascot Z score).
S.C: sequence coverage, percentage of amino acids sequence covered by matching peptides.
Theoretical molecular mass (Mr) and pI of matched sequence. Theor.: theoretical; pI, isoelectric point.
Estimated molecular mass (Mr) and pI of protein spot from the gel by ImageMaster software. Esti.: estimated.
Functional class codes according to Listilist Listeria genome database (
Expression in Δ1771 cells: + , protein spot significantly (p < 0.05) increased in Δ1771; − , protein spot significantly (p < 0.05) reduced in Δ1771.
Functional grouping of differentially expressed genes in Δ1771
A high-density NimbleGene microarray constructed for L. monocytogenes strain 4b F2365 (containing probes for 2821 genes) was employed to characterize global differential gene expression in the deletion mutant. In this analysis, 49 genes (including lm.G_1771) showed differential expression at the mRNA level with the threshold of a minimum twofold difference (p < 0.05). These genes could be classified into 8 functional categories (Table 3). It was found that the number of genes identified as differentially regulated using microarrays was larger than that of proteins identified in the proteomic study. This result is not unexpected, as the pH range of the IPG strips used was pH 4–7 and the pI of many proteins was out of this range.
Indicates the correlation between genes identified by microarray and the corresponding protein according to the proteomic analysis of the Δ1771 mutant.
Evaluation of differentially expressed genes by real-time quantitative PCR
Ten genes (dltA to dltD, srtA, sod, usp, gntR, LMOf2365_2147 and 2148) that were differentially regulated were confirmed by real-time quantitative PCR with normalizing to 16S rRNA gene (Fig. 3). Relative quantitative values were obtained using the comparative threshold cycle method (ΔΔCT ). The relative expression of the genes was determined in quadruplicate from total RNA extracted from three independent cultures. The resulting ratios were log2 values transformed and plotted against the log2 values transformed from microarray hybridization analysis. It was found that relative changes in gene expression obtained from microarray and real-time quantitative PCR experiments correlated in a linear relationship for each gene with r > 0.9, which was considered the threshold for strong correlation.
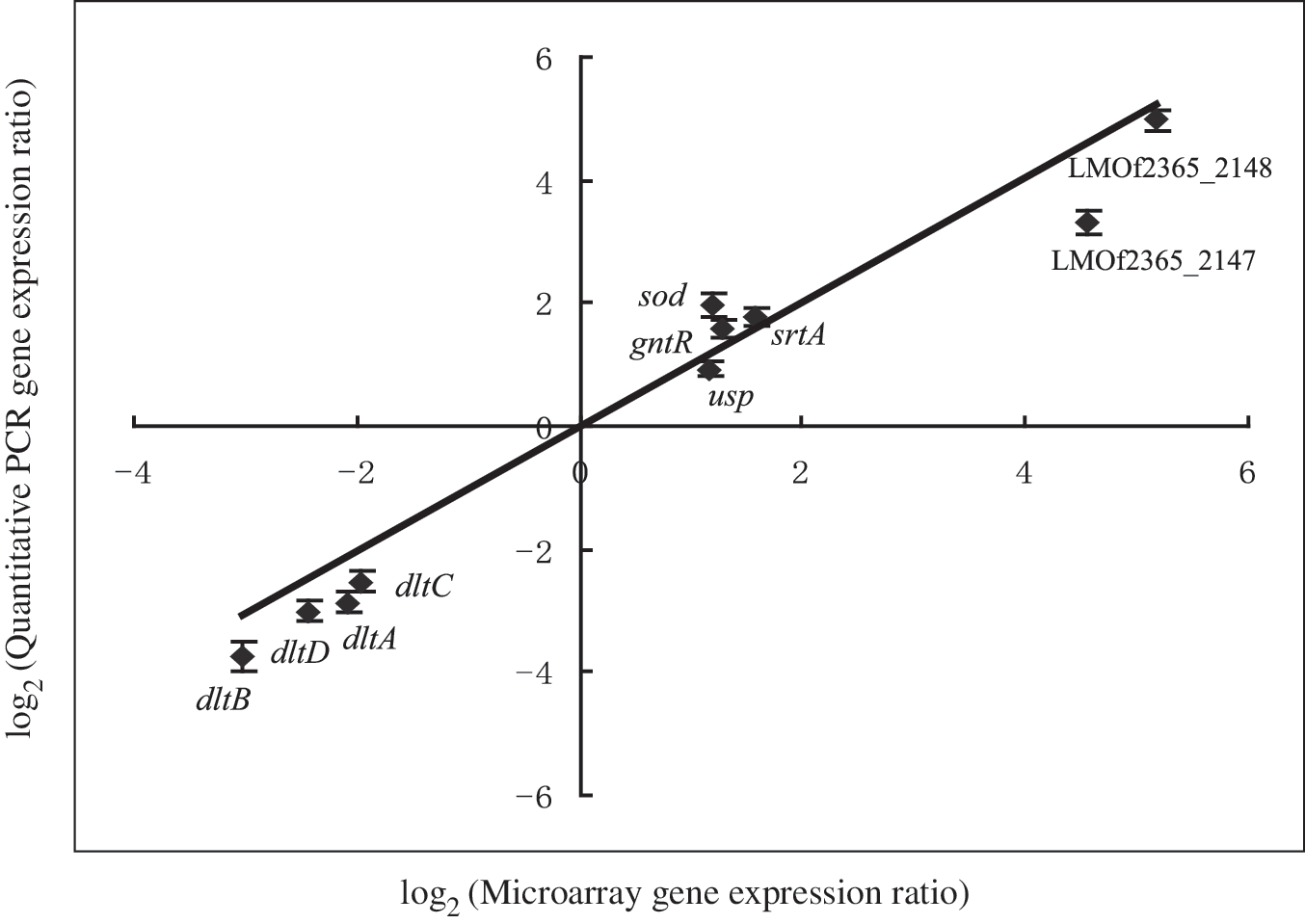
Comparison between differential responses in gene expression in L. monocytogenes measured by real-time quantitative PCR and microarray data from RNAs obtained from the wild-type and Δ1771 strain. PCR, polymerase chain reaction.
Discussion
An lm.G_1771 deletion mutant (Δ1771) that showed the same increased ability to form biofilms as LM-49 was constructed for further study. Several genes or proteins were found to be possibly influenced by lm.G_1771 by proteomic and transcriptomic analyses in this work. Among the identified genes, several encoded for cell surface proteins. Therefore, lm.G_1771 could participate in a novel signal transduction pathway that regulates the expression of the identified cell wall surface proteins. Some valuable information regarding this potentially novel signal transduction pathway may have been lost in this study, as many proteins had pIs beyond the tested range. However, the data from microarray analyses suggested that the lm.G_1771 could participate in a novel signal transduction pathway (Lm.G_1771 does not export AI-2, data not shown).
Three Listeria cell surface proteins (Lsps, LMOf2365_0347, 0543 and 1974) exhibited more than twofold greater expression in the mutant, and they likely function as drivers of biofilm formation in L. monocytogenes (Table 3). The characterization of some cell surface proteins in other bacteria has already demonstrated that they play important roles in biofilm formation. For example, the Enterococcus faecalis Esp cell surface protein was experimentally shown to be an important mediator of bacterial biofilm formation on a polystyrene surface (Toledo-Arana et al., 2001). An up-regulated Lsp LMOf2365_1974 protein showed 39% amino acid sequence similarity to Esp cell surface protein, and its role in biofilm formation by L. monocytogenes is still unknown. Further, these three Lsps harbor a C-terminal sorting signal with an LPXTG motif, covalently linked to the cell wall peptidoglycan by a transamidase named sortase (Mazmanian et al., 1999). Three other cell wall modifying enzymes (LMOf2365_0950, 1246 and 2741) were also expressed at a higher level in the mutant (Table 3). LMOf2365_0950 protein is a sortase A (SrtA) homolog in L. monocytogenes, as this protein shows 54% amino acid sequence similarity with SrtA from Streptococcus mutans (Levesque et al., 2005), which suggests that this SrtA homolog could have a similar role in biofilm formation by translocation of the three Lsps with a LPXTG motif to cell surfaces. It would be very interesting to study whether these cell surface proteins function individually at different steps or they work in concert to mediate biofilm formation.
From the microarray data, we also observed that the expression of the dlt operon is down regulated in the Δ1771 mutant. Genes in the dltA-dltD operon are known to be involved the incorporation of positively charged D-alanine residues into lipoteichoic acids (LTAs). Reducing alanyl LTAs should increase the negative surface charges on bacterial cells (Heptinstall et al., 1970) and the ester content determining the number of anionic sites on LTAs for autolysin binding (Wecke et al., 1997). Therefore, this may explain why the Δ1771 mutant is more sensitive to cationic antibiotics and more easily lysed in Triton X-100 compared with the wild-type strain (data not shown). Alanylation of LTAs also affects biofilm formation in other bacteria. In S. aureus, inactivating a Dlt enzyme by gene knockout abrogates bacterial adhesion to plastic (Gross et al., 2001), thereby negatively regulating biofilm formation. By contrast, a Lactobacillus rhamnosus dltD mutant exhibits a biofilm positive phenotype (Lebeer et al., 2007). In the current study, reduced expression was observed for all dlt genes in the mutant by microarray data and was further confirmed by real-time quantitative PCR (Fig. 3), which suggests a reverse relation between dlt gene expression and biofilm formation in L. monocytogenes.
Genes encoding the putative transcription factors that were down-regulated in the lm.G_1771 deletion mutant were identified. One is the GntR-like transcription factor that has been shown to regulate biofilm formation in E. faecalis (Ballering et al., 2009), and the other is MarR-like transcription factor. Therefore, functional analyses of the identified transcriptional factors and the cell wall surface proteins will yield important insights into the yet unknown signal transduction pathway involving lm.G_1771 and biofilm formation in L. monocytogenes. Further studies are needed to identify signaling molecules and signal transduction pathways that regulate biofilm formation in L. monocytogenes.
Footnotes
Acknowledgments
This work was jointly supported by the grant No. 30972485 and U1031003 from National Natural Science Foundation of China, the grants No. 2009BADB9B01 and No. 2009BAK43B31 from the Ministry of Science and Technology of China, and No. 08142200700, 2009DFA31770 and 09DZ0503300 from Science and Technology Commission of Shanghai Municipality to X. Shi; Danish Free Research Councils grants 274-07-0116 (FTP) and 272-05-400 (FNU) to Q. She, and grant 274-05-0073 (FTP) to S. Knøchel. The authors thank Andrea Maria Lorentzen at University of Southern Denmark for assistance with the protein identifications.
Disclosure Statement
No competing financial interests exist.