
Abstract
Outbreaks associated with leafy greens have focused attention on the transfer of human pathogens to these commodities during harvest with commercial equipment. Attachment of Escherichia coli O157:H7 on new or rusty spinach harvester blades immersed in spinach extract or 10% tryptic soy broth (TSB) was investigated. Bacteriophages specific for E. coli O157:H7 were evaluated to kill cells attached to blade. A cocktail of five nalidixic acid-resistant E. coli O157:H7 isolates was transferred to 25 mL of spinach extract or 10% TSB. A piece of sterilized spinach harvester blade (2 × 1") was placed in above spinach extract or 10% TSB and incubated at room (22°C) or dynamic (30°C day, 20°C night) temperatures. E. coli O157:H7 populations attached to blade during incubation in spinach extract or 10% TSB were determined. When inoculated at 1 log CFU/mL, E. coli O157:H7 attachment to blades after 24 and 48 h incubation at dynamic temperature (6.09 and 6.37 log CFU/mL) was significantly higher than when incubated at 22°C (4.84 and 5.68 log CFU/mL), respectively. After 48 h incubation, two blades were sprayed on each side with a cocktail of E. coli O157–specific bacteriophages before scraping the blade, and subsequent plating on Sorbitol MacConkey media–nalidixic acid. Application of bacteriophages reduced E. coli O157:H7 populations by 4.5 log CFU on blades after 2 h of phage treatment. Our study demonstrates that E. coli O157:H7 can attach to and proliferate on spinach harvester blades under static and dynamic temperature conditions, and bacteriophages are able to reduce E. coli O157:H7 populations adhered to blades.
Introduction
Contamination of fresh produce can occur anywhere throughout the production process. The use of contaminated manure can increase the presence of pathogenic microorganisms on produce (Barak and Liang, 2008). E. coli O157:H7 can persist on produce when irrigated with contaminated water (Solomon et al., 2003; Islam et al., 2004). Contaminated equipment used in harvesting may be able to transfer pathogens to fresh or fresh-cut produce. Mechanical spinach harvester blades that function close to the ground may come into contact with fecal matter from wild animals or improperly composted manure that can introduce E. coli O157:H7 to these blades.
During spinach harvesting, extracts from the cut spinach may provide a growth medium for pathogenic bacteria attached to the harvester blade, and present an opportunity for biofilm formation on the blade surface. Bacterial attachment and potential biofilm formation on harvester blades is a concern because sessile bacteria within biofilms will not be killed by treatment with chlorinated water.
Bacteriophages are widely distributed in the environment and represent part of the natural microbiological flora of foods. Phages are extremely specific to bacterial hosts, and their application against bacterial pathogens in food has been previously reported (Rees and Dodd, 2006). However, no information is available regarding the use of phages to control E. coli O157:H7 on harvesting equipment. We studied the attachment of E. coli O157:H7 on spinach harvester blades at constant and dynamic simulated field temperatures. The ability of phages to reduce E. coli O157:H7 on harvester blades was also evaluated.
Materials and Methods
Preparation of bacterial strains
Nalidixic acid (NA)-resistant E. coli O157:H7 strains RM 4406, RM 4688, and RM 1918 (clinical isolates from lettuce outbreaks), RM 4407 (clinical isolate from spinach outbreak), and RM 5279 (clinical isolate from bagged vegetable) were used in the study. Frozen cultures were initially thawed at room temperature (∼22°C) for 15 min, transferred into tryptic soy broth (TSB) containing 50 mg/L NA (Sigma-Aldrich), and incubated at 37°C for 24 h.
Bacteria were harvested from the 24-h culture by centrifugation (5000 g, 15 min), washed with phosphate-buffered saline (PBS, pH 7.2) twice, and adjusted OD600 of 1 that equates to 9 log CFU/mL. A cocktail of the five E. coli O157:H7 strains described above was prepared using an equal amount of each culture for inoculation in spinach extract and 10% TSB. The cell populations of individual strains were verified by the spot plate procedure using TSA supplemented with 50 mg/L NA.
Inoculation of harvester blades
A spinach extract was prepared by pummeling 25 g fresh spinach leaves (Spinacia oleracea) for 2 min in filtered stomacher bags (Fisher Scientific) using a laboratory blender (Bagmixer; Interscience) with 10 × (w:w) volume of 0.1% peptone water (Becton Dickinson). Twenty-five milliliters of spinach extract or 25 mL of 10% TSB in 50-mL conical tubes was inoculated separately with 250 μL of the five-strain E. coli O157:H7 cocktail to obtain a final population of 1 (low) and 4 (high) log CFU/mL. Two types of spinach harvester blades, new and rusty (used previously for spinach harvesting), obtained from a spinach farmer in Maryland were used. A sterilized spinach harvester blade (5 × 2.5 cm) was placed aseptically in inoculated spinach extract or 10% TSB in 50-mL centrifuge tubes and incubated at a static temperature (room temp, 22°C) or at a dynamic temperature (30°C 12 h, 20°C 12 h), simulating California day–night temperatures, in a growth chamber for up to 48 h. At 1, 24, and 48 h of incubation, two blades at each inoculum level were removed from tubes to determine bacterial populations adhered to blade.
Microtiter plate assay for biofilm
Overnight cultures were diluted 1:10,000 in spinach extract, TSB, or 10% TSB, and 200 μL was placed in sterile 96-well polystyrene microtiter plates (Fisher Scientific), and incubated under static conditions at 30°C for 48 h. The microtiter plate assay was performed using the procedure described by Patel and Sharma (2010).
Microbial analysis
Blades were removed from spinach extract or 10% TSB with a sterile forceps, gently rinsed in a10 mL sterile distilled water, and then transferred to a sterile Petri dish containing 10 mL PBS. A sterile Teflon “policeman” scraper (Fisher Scientific) was used to remove the bacterial mass from the blade by scraping each side for 1 min while the blade was immersed in PBS, and suspensions containing E. coli O157:H7 were serially diluted and spiral plated on Sorbitol MacConkey media (Acumedia) supplemented with 50 μg/mL NA (Sigma-Aldrich). Sorbitol-negative E. coli O157:H7 colonies were counted after 24 h incubation at 37°C. Randomly selected 8–10 colonies were confirmed using latex agglutination assay (Remel). Similarly, E. coli O157:H7 populations in spinach extract or 10% TSB were also determined at 1, 24, and 48 h.
Bacteriophage application
Bacteriophages specific for E. coli O157 previously collected and isolated from feedlot cattle feces (Callaway et al., 2006) were used in the study. The titer of the bacteriophage stock suspensions (n = 6 isolates) was determined by soft agar overlay technique using E. coli O157:H7 strains listed above. Four blades incubated for 48 h in spinach extract containing E. coli O157:H7 were used for phage treatment. Blades were gently rinsed in a 10 mL sterile distilled water and allowed to air-dry for 30 min. A hand-held air-brush (Badger Air-Brush Co.) filled with 20 mL of phage cocktail (8 log PFU/mL) was used to spray each side of the blade with 50 μL. Two blades sprayed with PBS served as a control. Treated blades were held at room temperature (22°C) in sterile Petri dishes. E. coli O157:H7 populations on control and bacteriophage-treated blades were analyzed after 2 h using the procedure as described above.
Statistical analysis
Two blades were analyzed at 1, 24, or 48 h to determine the effect of growth media or incubation temperature on E. coli O157:H7 on harvester blades. Four blades were used at times of analysis to determine the effectiveness of the bacteriophage in reducing E. coli O157:H7 populations. The data obtained from four replicates were analyzed by a 3 × 2 factorial analysis of variance (SAS 8.2) to determine the effects of growth media, incubation temperatures, and blade conditions using the Shaffer-R simulation method. In all cases, the level of statistical significance was p < 0.05.
Results
Recovery of E. coli O157:H7 populations in spinach extract and 10% TSB
E. coli O157:H7 populations recovered from spinach extract and 10% TSB incubated at room and dynamic temperatures are shown in Table 1. Initial E. coli O157:H7 populations were ∼1 log CFU/mL at low inoculation level. Under dynamic incubation temperature conditions (30°C 12 h, 20°C 12 h), E. coli O157:H7 populations in inocula recovered from spinach extract (6.07 log CFU/mL) and in 10% TSB (7.02 log CFU/mL) after 24 h were significantly higher than the corresponding E. coli O157:H7 populations recovered in spinach extract (4.10 log CFU/mL) and in 10% TSB (5.22 log CFU/mL) incubated at room temperature. E. coli O157:H7 populations increased with incubation time of 48 h in both spinach extract and 10% TSB. The recovery of E. coli O157:H7 in spinach extract (6.10 log CFU/mL) after 48 h was significantly lower than the E. coli O157:H7 populations recovered from 10% TSB (7.14 log CFU/g). The condition (new or rusty) of blades in spinach extract or 10% TSB did not affect (p > 0.05) the recovery of E. coli O157:H7 from spinach extract or 10% TSB at both dynamic and static temperatures (data not shown). E. coli O157:H7 populations recovered from spinach extract (6.36 log CFU/mL) and in 10% TSB (7.98 log CFU/mL) containing rusty blades that were incubated for 24 h at dynamic temperature were significantly higher than the corresponding E. coli O157:H7 populations in spinach extract (5.32 log CFU/mL) and 10% TSB (5.52 log CFU/mL) incubated at room temperature (data not shown). At high inoculum levels, E. coli O157:H7 recovered from spinach extract (7.99 log CFU/mL) or 10% TSB (7.99 log CFU/mL) after 24 h at dynamic temperature incubation were similar (p > 0.05) to the E. coli O157:H7 detected in spinach extract (7.36 log CFU/mL) or 10% TSB (7.08 log CFU/mL) at room temperature incubation.
Spinach extract and 10% TSB were incubated at dynamic (30°C day, 20°C night) or static (22°C) temperatures.
Means with different letters in the row are significantly different (p < 0.05).
Means with different letters in the columun within the incubation time are significantly different (p < 0.05).
TSB, tryptic soy broth.
Attachment of E. coli O157:H7 on spinach harvester blades
E. coli O157:H7 populations recovered from new and rusty spinach harvester blades incubated for 1 h in spinach extract or 10% TSB were ∼0.5–0.9 log CFU/blade (Fig. 1A, B) when inoculated at low level. E. coli O157:H7 populations recovered from new blades were higher (p > 0.05, 0.77–0.94 log CFU/blade) than populations recovered from rusty blades (0.54–0.66 log CFU/blade). Recovery of E. coli O157:H7 from new blades incubated at dynamic temperature for 24 h in spinach extract and 10% TSB (4.69 and 5.89 log CFU/blade) was significantly higher than those incubated at room temperature in spinach extract and 10% TSB (4.04 and 2.57 log CFU/blade), respectively. Likewise, E. coli O157:H7 populations recovered from rusty blades incubated at dynamic temperature in spinach extract and TSB (5.47 and 5.48 log CFU/blade) for 24 h were significantly higher (p < 0.05) than those incubated at room temperature in spinach extract and TSB (4.31 and 3.67 log CFU/blade), respectively. E. coli O157:H7 attachment to blades increased with an increase in incubation to 48 h in spinach extract or 10% TSB.
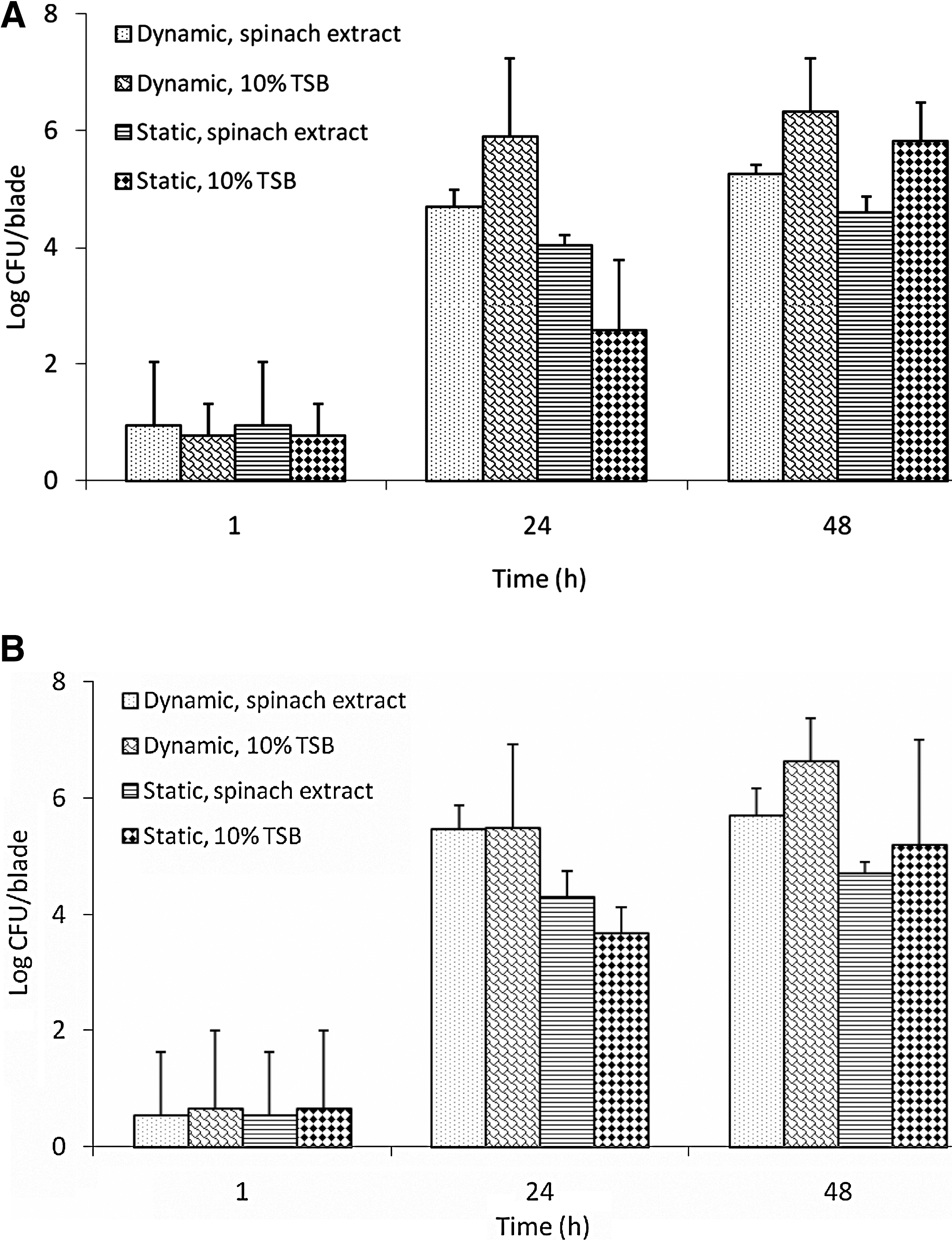
Adherence of Escherichia coli O157:H7 to
At the high inoculation level, adherence of E. coli O157:H7 on harvester blades ranged from 2.40 to 3.06 log CFU/blade after 1 h of incubation (Fig. 2A, B). At least 6 log CFU/blade of E. coli O157:H7 were recovered from blades in spinach extract or 10% TSB after 24 h incubation at either static or dynamic temperatures. E. coli O157:H7 populations recovered from new blades incubated in 10% TSB for 24 h at dynamic temperature (7.05 log CFU/blade) were significantly higher than the populations recovered from blades incubated in 10% TSB at room temperature (6.00 log CFU/blade). Populations of bacteria adhered to blade incubated in spinach extract or 10% TSB for 48 h were not different (p > 0.05) from the corresponding populations detected from blade after 24 h incubation.
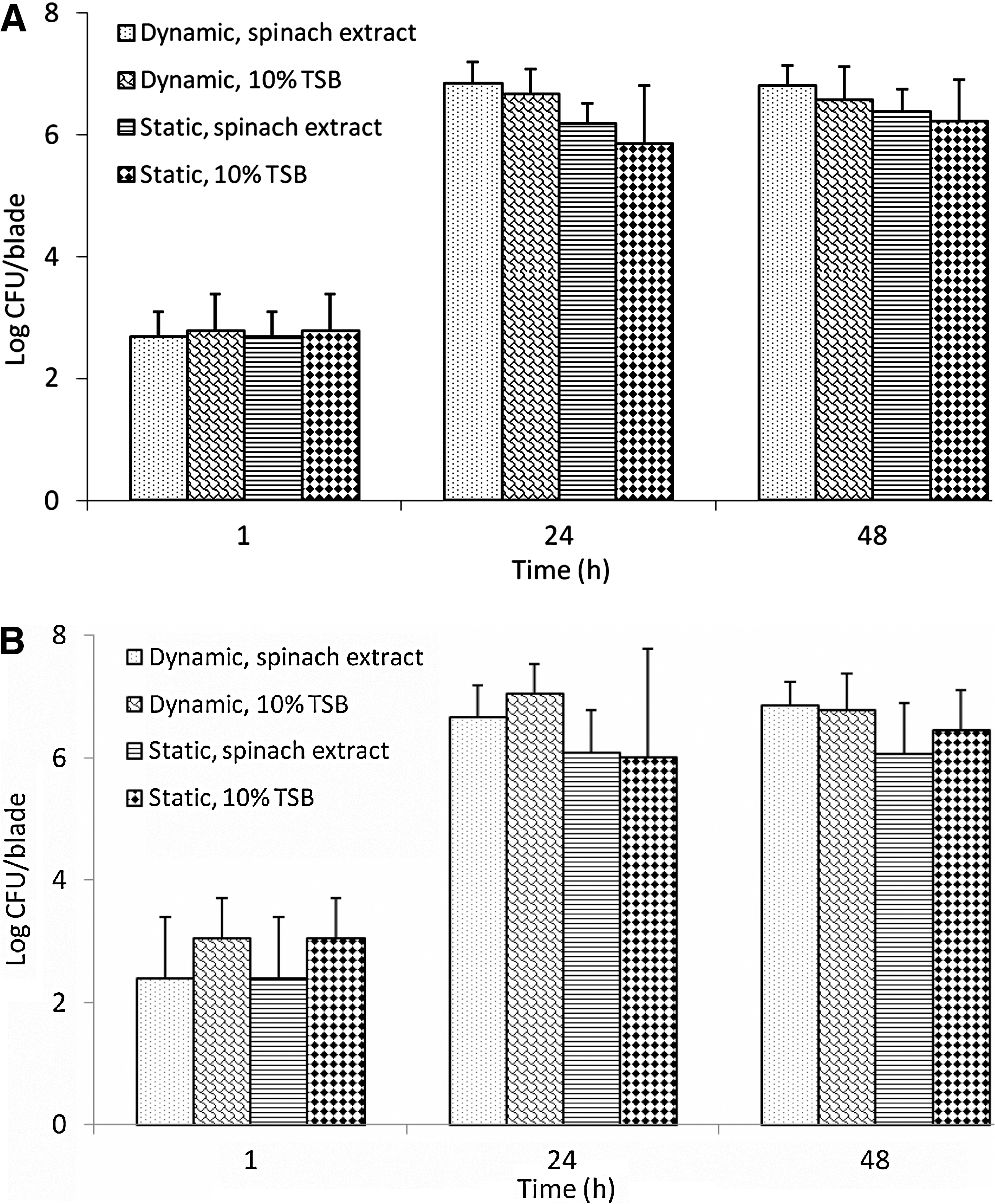
Adherence of E. coli O157:H7 on
Biofilm formation on polystyrene microtiter plate
The biofilm formation of E. coli O157:H7 strains determined by microtiter well plate assay is shown in Figure 3. The strength of biofilm was determined using method described by Stepanovic et al. (2004). Biofilm formation was significantly affected by the strain and the growth medium used in microtiter plate assay. E. coli O157:H7 strain 5279 was the strongest biofilm former (0.38, p < 0.05) followed by strain 4688 (0.33) and strain 1918 (0.31) when grown in 10% TSB. Biofilm formed by strains 4406 (0.22) and 4407 (0.25) in 10% TSB were similar (p > 0.05) to the biofilm formed with uninoculated control. Biofilm formed by all E. coli O157:H7 strains in 10% TSB with the exception of strain 4406 were significantly higher than the corresponding biofilm formed in full strength TSB. Biofilm formed by E. coli O157:H7 strains in TSB ranged from 0.16 to 0.20 and were not significantly different (p > 0.05).
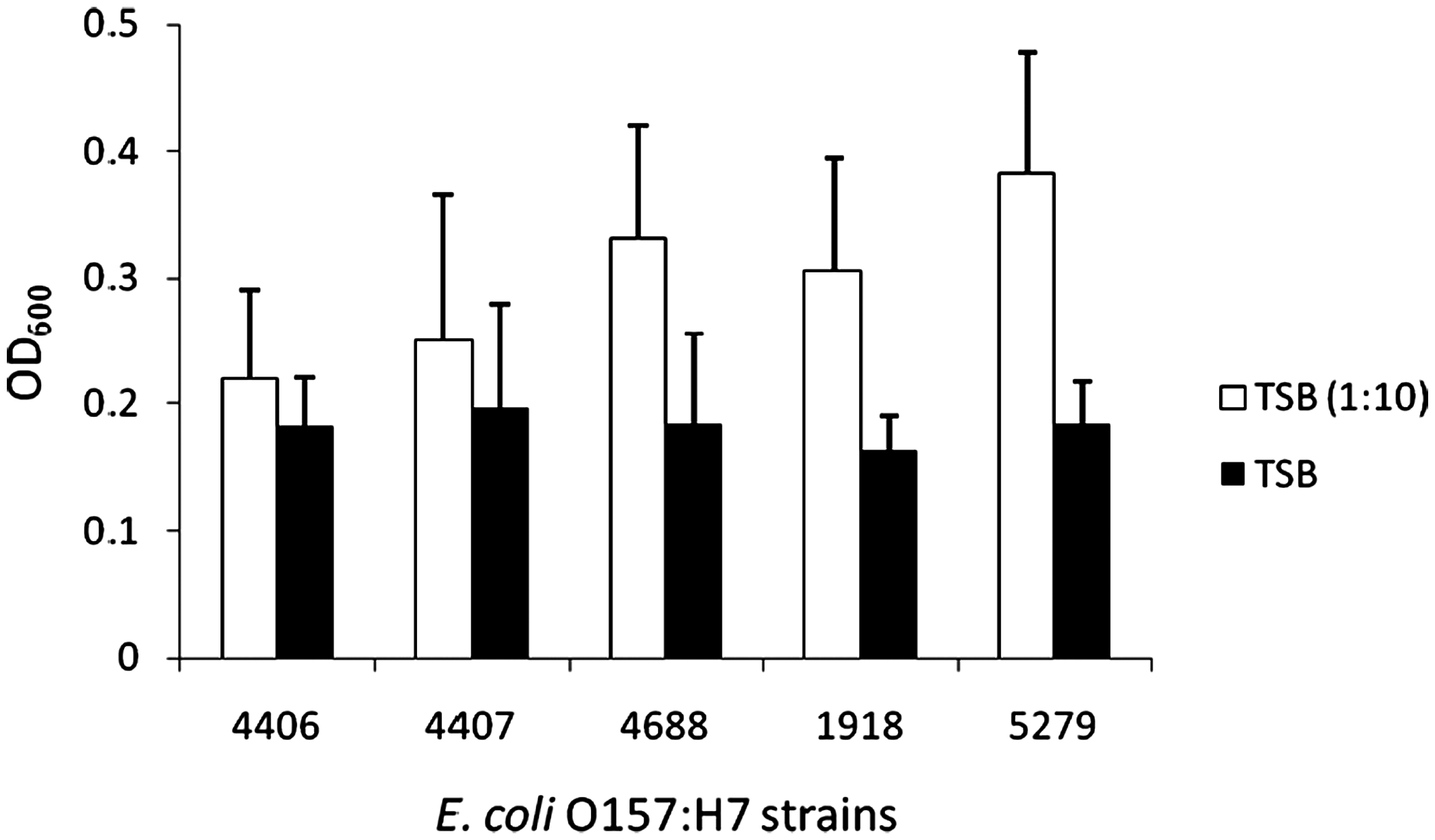
Biofilm formation by E. coli O157:H7 strains in full-strength TSB and 10% TSB as determined by crystal violet binding assay. The results (OD600 values) are the means of 24 wells. The error bars represent standard deviation.
Effect of bacteriophage on attached E. coli O157:H7 cells
Initial E. coli O157:H7 populations attached to harvester blade were 4.87 log CFU/blade (data not shown). The populations of E. coli O157:H7 attached to blade were undetectable (<0.4 log CFU) after 2 h exposure of phage treatment. E. coli populations detected on blade following 2 h control (PBS) treatment (4.40 log CFU) were not significantly different from the initial E. coli O157:H7 populations (4.87 log CFU) on blade.
Discussion
The work described in our study shows that E. coli O157:H7 cells can attach to spinach harvester blades under both static and dynamic temperature conditions. Further, spinach extract has shown to provide ample nutrients for attached E. coli O157:H7 cells to grow on blade surfaces. Although many different factors are involved in bacterial colonization and biofilm formation on the equipment or produce surface, the type of surface might affect biofilm formation. Curli, a thin, coiled fimbriae-like extracellular polysaccharide, produced by E. coli O157:H7, enhanced biofilm formation on stainless steel surfaces (Ryu and Beuchat, 2005, Silagyi et al., 2009). These authors found significant transfer of E. coli O157:H7 from stainless steel surface to various produce surfaces. We found biofilm formation in microtiter plate assay when E. coli O157:H7 strains were grown in 10% TSB. Our results are in agreement with other studies that found variable biofilm formation with nutrient availability and type of broth (Ryu et al., 2004; Silagyi et al., 2009). The stronger biofilm in diluted TSB medium could be attributed to the induction of biofilm under starvation stress (Stepanovic et al., 2004). Biofilm formation on polystyrene microtiter plate and harvester blade could be different as surface properties (hydrophobic vs. hydrophilic) will also influence bacterial attachment and subsequent biofilm formation. Absorbance readings for E. coli O157:H7 in 10% spinach extract were inconclusive (data not shown). The presence of organic matter in spinach extracts might have hindered the accurate detection of crystal violet stain attached to biofilm in the wells of the microplate.
Cut spinach can release nutrients in the extracts which bacterial pathogens can utilize for growth and promote attachment to harvesting equipment. A study evaluated the effect of field coring for survival and growth of E. coli O157:H7 on precored lettuce (McEvoy et al., 2009). A coring knife, used to harvest fresh-cut iceberg lettuce, was inoculated with ∼5 log CFU of E. coli O157:H7 and was able to contaminate at least nine heads of lettuce. Similarly, our study shows that spinach extracts on harvester blades support the adherence and growth of E. coli O157:H7 at static and dynamic temperatures, and could potentially cross contaminate spinach during harvesting. Neither the condition of the blades nor growth medium significantly influenced adherence of E. coli O157:H7 on blade, indicating that spinach extract is as rich as 10% TSB in supporting the growth and adherence of E. coli O157:H7 on harvester blade. It also indicates that E. coli O157:H7 are not more able to adhere to rusty blades than new ones.
Spinach harvester blades are commonly sanitized with hypochlorite solutions. The presence of organic matter such as spinach extracts and soil could reduce the free chlorine available for effective killing of E. coli O157:H7 on harvester blades. Treatment at 200 μg/mL total hypochlorite did not eliminate E. coli O157:H7 biofilms on stainless steel surfaces (Ryu and Beuchat, 2005), and total hypochlorite concentrations of 25–200 mg/L failed to completely remove biofilm formed by E. coli O157:H7 or Salmonella from stainless steel surfaces (Ueda and Kuwabara, 2007). Limited efficacy of hypochlorite solutions in the presence of organic matter on stainless steel surfaces has led to the evaluation of alternative treatments for harvester blade sanitation. We were able to achieve ∼4 log reduction of E. coli O157:H7 on blades after 2 h of bacteriophage spray treatment. Individual phage suspensions were able to kill five E. coli O157:H7 strains used in the study. However, a cocktail of phage suspensions was used to overcome possible generation of resistant mutants. The spray application of the bacteriophage is compatible with the sanitation practices for harvesting equipment and therefore may provide a practical alternative to dipping, brushing, or sponging on a chlorine solution.
Our study indicates that E. coli O157:H7 can attach to spinach harvester blades under static and dynamic temperature conditions when spinach extract is in contact with the blade. The potential cross-contamination of fresh-cut spinach by E. coli O157:H7 during harvesting deserves evaluation as a risk factor in produce safety. Bacteriophages specific for E. coli O157:H7 may provide a potential alternative treatment to hypochlorite. This is the first report on the effect of bacteriophages against E. coli O157:H7 on harvester blades. Further study on biofilm formation on spinach harvester blades is required to reduce the risk of pathogenic bacterial contamination during harvesting of fresh produce.
Footnotes
Acknowledgments
Authors thank Dr. Bryan Vinyard for statistical analysis. We also thank Sheena Crutchfield, Shanta Adeeb, and Katherine Darlington for the laboratory assistance with this study.
Disclosure Statement
No competing financial interests exist.