
Abstract
One QnrA1-producing Klebsiella pneumoniae isolate GDKA1 from chicken was detected. The qnrA1 gene on plasmid pGDKA1 was located in a genetic environment similar to that in In36 on plasmid pHSH1 and could be cotransferred to Escherichia coli J53 AzR with other resistances by a conjugation experiment. Upstream of the qnrA1 gene, there was a class I integron with the dfrA27 and aadA2 cassettes. Similar genetic environments of qnrA1 in Enterobacteriaceae isolates from both human and animal origin might, to some extent, demonstrate similar mechanisms of qnrA distribution. The presence of qnrA1 in health animal commensal bacteria should be worthy of note. This is the first report of qnrA1 in K. pneumoniae and dfrA27 in an Enterobacteriaceae isolate of animal origin.
Introduction
One (0.51%) qnrA1-carrying K. pneumoniae isolate, strain GDKA1 was detected in a total of 195 Enterobacteriaceae isolates from healthy chicken in five Guangdong farms in China in 2007. In this study, we characterized the isolate and the genetic environment of qnrA1.
Materials and Methods
Bacterial strains
GDKA1 is a qnrA1-carrying K. pneumoniae strain isolated from a random rectal swab sample of chicken in a farm located in Guangdong China in 2007. Escherichia coli J53 AzR was used as recipient cell in conjugation experiment. E. coli DH5α served as recipient strain for DNA cloning. Strains were routinely grown at 37°C in Luria-Bertani medium except as noted otherwise.
Susceptibility testing
Minimum inhibitory concentration (MIC) for eight antimicrobials belonging to different classes or subclasses: nalidixic acid, ampicillin, chloramphenicol, cefotaxime, tetracycline, ciprofloxacin, sulfamethoxazole, and spectinomycin were determined by E-test strips (AB Biodisk).
Conjugation experiment
Conjugation was performed using E. coli J53 AzR as the recipient strain. To avoid unintentional selection of quinolone-resistance mutations in the recipient, transconjugants were selected on Trypticase soy agar plates containing sodium azide (100 μg/mL) and sulfamethoxazole (200 μg/mL). A transconjugant TGDKA1 was selected, and the presence of the qnrA1 gene was confirmed by polymerase chain reaction (PCR).
Plasmid manipulations and sequencing
Plasmid pGDKA1 was isolated from TGDKA1 using Qiagen Midi Kit (Qiagen) and digested with EcoRI (Takara). DNA fragments were cloned into vector pUC19 (Takara) digested with the same enzyme. Recombinants were transformed into competent E. coli DH5α by heat-shock method. A transformant carrying qnrA1 was identified by PCR. Recombinant plasmids were extracted with QIAGENPrep Plasmid Midi Kit (Qiagen), and the nucleotide sequence of the insert carrying qnrA1 was determined with specific primers on an ABIPRISM 310 Genetic Analyzer (Applied Biosystems).
PCR amplification and sequencing
Long-accurate PCR by LA Taq (Takara) using primers targeting the class I integrase encoding intI1 gene (5′-AACAAACGATGCTCGCCTTCCA-3′) and the qnrA1 gene (5′-TGAGAAATCCTCTTGCTGA-3′) on pGDKA1 was performed on an iCycler thermal cycler (Eppendof) as follows: initial denaturation step of 5 min at 94°C, 32 cycles consisting of 94°C for 45 sec, 48°C for 45 sec, and 72°C for 5 min and then a final extension step of 10 min at 72°C. PCR-amplified fragments were purified with the QIAquick PCR Purification Kit (Qiagen), then cloned into the pMD19-T simple vector (Takara), and finally transformed into competent E. coli DH5α as described earlier. Sequencing was carried out by walking with a series of specific primers (Takara).
Mutations in quinolone resistance determining regions of gyrA, gyrB, parC, and parE were detected by PCR and sequencing as previously described (Yue et al., 2008).
DNA analysis
All of the DNA sequences obtained were compared with relevant sequences in the GenBank database by using the BLAST algorithm (
Nucleotide sequence accession numbers
The partial sequence of plasmid pGDKA1 has been submitted to the GenBank database and assigned accession number EU722351.
Results and Discussion
Susceptibility testing showed that strain GDKA1 was resistant to ampicillin, sulfamethoxazole, spectinomycin, chloramphenicol, and tetracycline (Table 1). In contrast, this strain was susceptible to cefotaxime (MIC, 0.047 μg/mL) and ciprofloxacin (MIC, 0.19 μg/mL) and intermediate to nalidixic acid (MIC, 12 μg/mL). A transconjugant TGDKA1 was selected during the mating experiment, and hybridization with qnrA1-specific probes revealed that the qnrA1 gene was located in pGDKA1 (data not shown). Tranconjugant TGDKA1 showed a little higher quinolone MICs (Table 1) than E. coli J53 AzR but not as high as to be defined as resistant according to the MIC susceptible breakpoint ≤1 mg/L and resistant breakpoint ≥4 mg/L for CLSI. This could be explained by the fact that qnrA1 gene just confers resistance to nalidixic acid and decreased suscpetibility to fluoroquinolones. The donor strain GDKA1 showed more resistance to nalidixic acid than its transconjugant TGDKA1, indicating that there must be some other mechanism employed by the donor. Sequencing of quinolone resistance determining regions revealed that the donor had an S83I mutation in gyrA, whereas the recipient, transconjugant strain did not contain mutations associated with quinolone resistance. Resistances to spectinomycin, ampicillin, tetracycline, and sulfamethoxazole were cotransferred (Table 1).
MIC, minimum inhibitory concentration; AM, ampicillin; CI, ciprofloxacin; CL, chloramphenicol; CT, cefotaxime; NA, nalidixic acid; SC, spectinomycin; SX, sulfamethoxazole; TC, tetracycline.
A 7, 522-bp continuous DNA sequence obtained by digestion of pGDKA1 with EcoRI revealed that similar to many other plasmids reported (Wang et al., 2003; Mammeri et al., 2005; Rodríguez-Martínez et al., 2007), qnrA1 was located in a sul1-type integron structure and was embedded downstream of an insertion sequence ISCR1 element. The quinolone resistance gene was located immediately upstream of a truncated version of qacEΔ1/sul1, which is identical to that identified in pQR-1 of E. coli Lo from a patient at Bicêtre Hospital (Mammeri et al., 2005). Downstream of the qacEΔ1/sul1 structure, insertion sequence IS26 was found between a kanamycin resistance aph(3')-I gene with a relic of the Tn1696-like transposase tnpA1696 gene and a truncated version of the samB gene with plasmid partition genes parB and parA.
Amplification experiments using primers targeting intI1 and qnrA1 detected an integron on pGDKA1. Upstream of qnrA1, this integron harbored two gene cassettes, dfrA27 and aadA2, encoding resistance to trimethoprim and aminoglycosides, respectively. dfrA27 was first reported from E. coli in China (Wei et al., 2009), and this is the first identification of dfrA27 in Enterobacteriaceae isolates of animal origin. DNA analysis showed that the dfrA27 and aadA2 cassettes were expressed from a weak version of Pc promoter region, and low level of transcription was speculated. Genes flanking qnrA1 in pGDKA1 and other representative plasmids are shown in Figure 1.
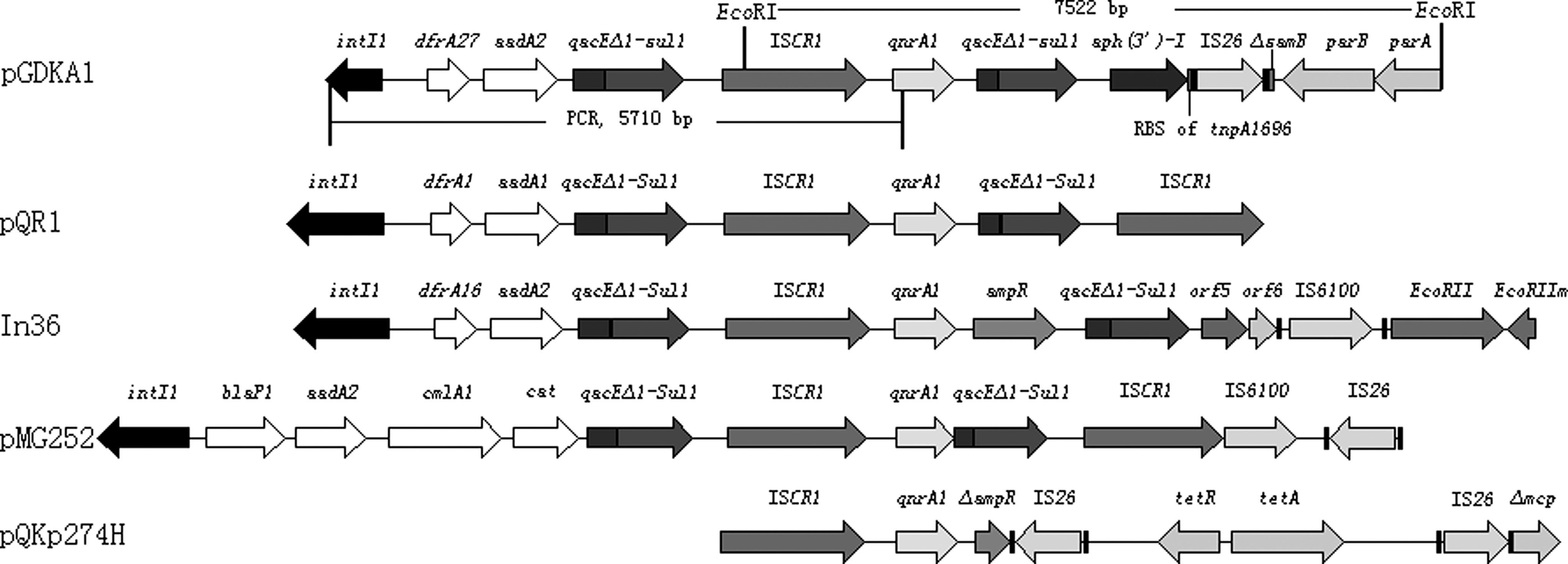
Genes flanking qnrA1 in pGDKA1 and other representative plasmids. Open reading frames are shown as arrows or horizontal boxes with an arrow indicating the orientation of the coding sequence with the gene name above the corresponding boxes. (GenBank accession numbers for pQR1, In36, pGM252, and pQKp274H (Lavilla et al., 2008) are AY655485, AY259085, DQ831140, and EF682136, respectively.)
To our knowledge, the qnrA1 determinants previously described for complex sul1-type integrons were all inserted between ISCR1 and a second copy of qacEΔ1/sul1 in the same orientation as the transposes of ISCR1 (Strahilevitz et al., 2009). In pGDKA1, the genetic environment of qnrA1 was generally homologous to In36 except for the gene cassettes in the upstream integron and absence of ampR and the downstream qacEΔ1, which was truncated. The same truncated qacEΔ1 downstream of qnrA1 was also identified in plasmid pQR1 (Mammeri et al., 2005). In contrast, the genes following the second copy of qacEΔ1/sul1 were variable. In pMG252 carried by a K. pneumoniae strain from Birmingham, AL (Jacoby, 2005), a second copy of ISCR1 was present, whereas in In36 on the plasmid pHSH1 of E. coli isolated from China (Wang et al., 2003), orf5 was identified instead. Further, many other resistance genes, truncated genes, gene relics, and mobile elements such as IS6100 and IS26 were often found in these genome regions, thus indicating frequent insertion and deletion events. Similar genetic environments of qnrA1 in Enterobacteriaceae isolates of both human and animal origin might demonstrate a similar target site of recombination events mediating incorporation of the quinolone resistance determinant. This could, to some extent, reflect homological mechanisms of qnrA1 distribution. Additionally, qnr genes providing low-level quinolone resistance facilitate the recovery of chromosome-encoded target mutants resulting in higher-level resistance (Martínez-Martínez et al., 1998), so they could at least partly be responsible for the increasing quinolone resistance in field strains.
In many Enterobacteriaceae strains, genes for antibiotic resistance and pathogenicity are usually plasmid borne. Thus, it will be a further threat to public health when such plasmids are conjugative (Lawley et al., 2004). Additionally, intestinal bacteria not only exchange resistance genes among themselves but might also cause bacteria passing through the colon to acquire and transmit antibiotic resistance genes (Salyers et al., 2004). Antibiotic resistant bacteria of animal origin may pass antibiotic resistance genes to bacteria that normally reside in humans (Salyers and Shoemaker, 2006). The present of qnrA1 on a conjugative plasmid of health animal commensal bacteria widened the dissemination of qnrA1 gene and should be worthy of note.
Footnotes
Acknowledgments
This study was supported in part by Chinese National Science Foundation and Guangdong Provincial United Foundation (U0631006), Provincial Science Foundation of Guangdong (5200638), the Program for Chang Jiang Scholars and Innovative Research Team in University (Grant No: IRT0723), and the 45th grant from China Post Doctor Science Foundation (20090450875). We thank George A. Jacoby, David C. Hooper (Division of Infectious Diseases, Massachusetts General Hospital), and Minggui Wang (Division of Infectious Diseases, Huashan Hospital, Fudan University) for kindly providing the E. coli strains J53AzR.
Disclosure Statement
No competing financial interests exist.