
Abstract
In nature the foodborne pathogen Listeria monocytogenes lives as a saprophyte where it can contaminate preharvest produce. This environment can present many stresses such as ultraviolet light, variations in temperature and humidity, and oxidative stress from growing plant matter in the soil. The alternative sigma factor Sigma B, encoded by sigB, controls the response to most stresses in L. monocytogenes. Fitness in soil and on radishes sown and grown in contaminated soil was measured in a wild-type and an isogenic sigB operon mutant strain to determine if the sigma factor was necessary for life in these niches. Levels of wild-type and mutant strains were monitored in contaminated soil over the course of radish gestation from seed to mature tuber, and levels on mature radishes were determined. The wild-type strain was able to survive in soil over the 4 weeks of the experiment at levels of 4–7 log CFU/g soil, and the levels of the sigB mutant were reduced by 1–2 log from the wild type. The mutant showed reduced levels in soil by 6 h after inoculation, which was partially recovered when the mutant was complemented, and stayed at a reduced level over the next 4 weeks. Upon harvest, 3–4 log CFU/g of wild-type L. monocytogenes was detected on radish surfaces, and the bacteria could not be washed off under running water. On mature radishes populations of the mutant strain were 1–2 log CFU/g lower than the wild type. The levels on mature radishes reflected the levels in the soil at 4 weeks. The conclusions are that the Sigma B operon is necessary for initial adaptation to the soil environment, and plays a role in maintaining the population, but does not play a role in attachment or colonization of the radish.
Introduction
Studies of the molecular mechanisms for L. monocytogenes attachment and colonization of plants indicate that flagella play a role, and the fitness of sprout attachment is strain dependent (Gorski et al., 2003, 2004, 2009). When exposed to plant tissue, this pathogen induces genes for carbohydrate, amino acid, and nucleic acid metabolism, and transport systems (Palumbo et al., 2005) similar to Pseudomonas and Rhizobium (Rainey, 1999; Lugtenberg et al., 2001).
One aspect important for bacterial colonization of plants is adaptation to stress. Escherichia coli O157:H7 induces genes important for resistance to oxidative stress when exposed to lysates of lettuce leaves (Kyle et al., 2010). The stress-response sigma factor rpoS is important for the attachment of Salmonella enterica serotype Newport to alfalfa sprouts (Barak et al., 2005). An rpoS mutant of Pseudomonas fluorescens has lower survival in the rhizosphere of bean plants in comparison to the wild type (Stockwell and Loper, 2005). L. monocytogenes sprout colonization is more efficient in strains with better resistance to oxidative stress (Gorski et al., 2008). The rpoS analog in L. monocytogenes, sigB, regulates responses to low pH, oxidizing conditions, starvation, and osmotic variations (Ferreira et al., 2001; Fraser et al., 2003). Rhizosphere growth requires dealing with variations in humidity, nutrients, and secretion of plant compounds into the soil. In this study we asked if growth and survival of a sigB mutant of L. monocytogenes was negatively affected in the rhizosphere of radish plants, and if the mutant was less likely to colonize radishes grown in the same soil.
Materials and Methods
Strains, plasmids, media, and culture conditions
L. monocytogenes RM2387, serotype 4b, was the wild-type strain (Gorski et al., 2003). RM4721, the sigB deletion strain, was constructed in the RM2387 background. Strains were typically grown on a rotary shaker at 100 rpm in TSYE medium (Bacto-Tryptic Soy Broth without Dextrose + 0.6% Bacto-Yeast Extract; Becton Dickinson) at 30°C for soil studies, and Brain Heart Infusion Medium (BHI; Difco) at 37°C for acid and oxidative stress studies. Modified Oxford Agar (MOX; Difco) was used to selectively isolate L. monocytogenes from inoculated soil and radishes. Mutants were constructed using pAUL-A, an L. monocytogenes shuttle vector with a temperature-sensitive origin of replication for gene replacement (Schaferkordt and Chakraborty, 1995), which was maintained with erythromycin (1 μg/mL) and lincomycin (25 μg/mL) in L. monocytogenes and erythromycin (300 μg/mL) in E. coli DH10B. The complementation plasmid in pAM401 was maintained in L. monocytogenes and E. coli with chloramphenicol (15 μg/mL) (Wirth et al., 1986). Phosphate-buffered saline (PBS) was a diluent and contained 150 mM NaCl and 10 mM sodium phosphate (pH 7.2).
SigB mutant construction
Primer sequences are in Table 1. The sigB gene from RM2387 was amplified from the genome using primers designed against the L. monocytogenes EGD-e genome sequence (
Restriction sites engineered into primer sequences are underlined.
A complementation plasmid was constructed in pAM401 (Wirth et al., 1986) with the sigB operon and promoter from RM2387. The resulting plasmid was electroporated into the sigB mutant. The control was the deletion strain carrying pAM401 without insert.
DNA sequence analysis
The sigB gene and promoter region from RM2387 were sequenced from three independently generated PCR amplicons. The deletion construct was also sequenced. Big Dye Terminator v. 3.1 (Applied Biosystems) was used on an ABI Prism 3130xl DNA Analyzer. Sequences were assembled with DNA Star (DNA Star).
Resistance to acid and oxidative stress
Strains were tested for their capacity to survive lethal acid exposure by measuring culture death upon exposure to BHI medium containing 100 mM acetic acid, adjusted to pH 2.5 with HCl as described (Ferreira et al., 2003). The resistance of the strains to oxidative stress by measuring the percent survival of a culture to survive 15 min of exposure to cumene hydroperoxide was done as described (Gorski et al., 2008). Both acid and oxidative stress experiment cultures were recovered on BHI agar.
Plant and soil studies
Supersoil (Scotts) was placed into 10-cm-diameter pots leaving 2 cm from the top free. Overnight L. monocytogenes cultures were diluted 1:10 in fresh TSYE and grown to mid-log phase. After pipetting 20 mL of a 106 CFU/mL aqueous L. monocytogenes suspension into each pot, they were incubated in random arrangements in a controlled-environment plant growth chamber with 11 h days (20°C, 4 UV, and 3 luminescent lights), 13 h nights (16°C, no light), and 70% humidity.
After 20–24 h the soil was sown with radish seeds (Rhafinus sativus cv. Cabernet; Johnny's Selected Seeds) that we treated to remove plant pathogens by washing in 70% ethanol for 5 min followed by two water rinses, a 10 min of incubation in 1% NaOCl, and three water rinses. Six seeds were sown per pot, and there were 6 replicate pots per strain. Pots with no seeds and with no L. monocytogones were controls. Immediately after sowing, the pots were given 30 mL of sterile, deionized water, the soil was sampled for the first time, and the pots returned to the growth chamber and watered regularly every 2–3 days. After 1 week the plants were thinned to three per pot.
Sampling of the soil consisted of transferring 1 g from the area surrounding the seed/tap root/tuber into a 15 mL sterile, conical tube. Ten milliliters of sterile H2O was added to the tube, which was vortexed for 10–20 s, the contents were allowed to settle for a minute, and the aqueous phase was diluted and plated onto MOX. Soil was sampled weekly.
After 4 weeks the radishes were harvested by pulling and removing both the green tops 1 cm from the top, and the tap root from the bottom, and weighed. One radish from each pot was wiped visibly free of soil with a Kimwipe, and placed in a Ziploc bag with 25 mL PBS. The second radish from each pot was rinsed under running water from a squeeze bottle for 15–20 s, rubbed with gloved hands (to mimic home washing under running water), and placed in a Ziploc bag with 25 mL of PBS. The radishes were rubbed in the bags for 1 min, and then the bags sonicated for 2 min in an Aquasonic Model 75T bath sonicator (VWR). The bagged radishes were rubbed again for 30 s, the PBS in the bag diluted, and plated onto MOX. The final radish from each pot was rinsed off under running water and placed in a beaker with 95% ethanol for 2–3 min, and then the ethanol was flamed off of the surface, before placing the radish into a stomacher bag with 25 mL of PBS. These radishes were homgenized in a Stomacher blender 1 min at normal speed, and the suspension was plated onto MOX. All MOX plates were incubated at 30°C for 1–2 days, and the characteristic Listeria colonies (bluish-white surrounded by black precipitate) were counted.
Statistical analysis
All experiments were performed at least four times with at least five replicates. All data shown are means with standard deviations or standard error. GraphPad InStat version 3.06 (GraphPad Software) was used for statistical analysis. Student's t-test was used for comparisons of two values, and one-way analysis of variance followed by Dunn's Multiple Comparisons test for comparison of more than two values.
Results
Construction and confirmation of the sigB deletion mutant
Sequencing and PCR confirmed that an internal 72% of the sigB gene (559 nucleotides) was deleted, leaving bases 1–99 and 658–780 intact. The deletion was out-of-frame, so expression of the downstream rsbX, a negative regulator of sigB, was tested. Reverse transcriptase–PCR showed a product from rsbX primers in the deletion strain, indicating that the rsbX ribosomal binding site was intact, but the functionality of RsbX was unknown (data not shown). The GenBank accession number for the sigB allele of RM2387 is DQ858351. The complemented mutant was confirmed by plasmid purification from the complemented mutant, and PCR verification that the strain contained both the deletion construct in the chromosome and the intact operon in the plasmid.
Stress resistance
Compared to wild type, the mutant exhibited 2–3 logs lower levels and was not detected after 90 min of acid exposure (Fig. 1). The complemented mutant showed recovery at 30 min (p > 0.05) and partial recovery thereafter. At 30 min the complemented mutant had higher levels than the empty vector strain (p < 0.001), but after that they were equivalent to each other and the mutant strain (p > 0.05).
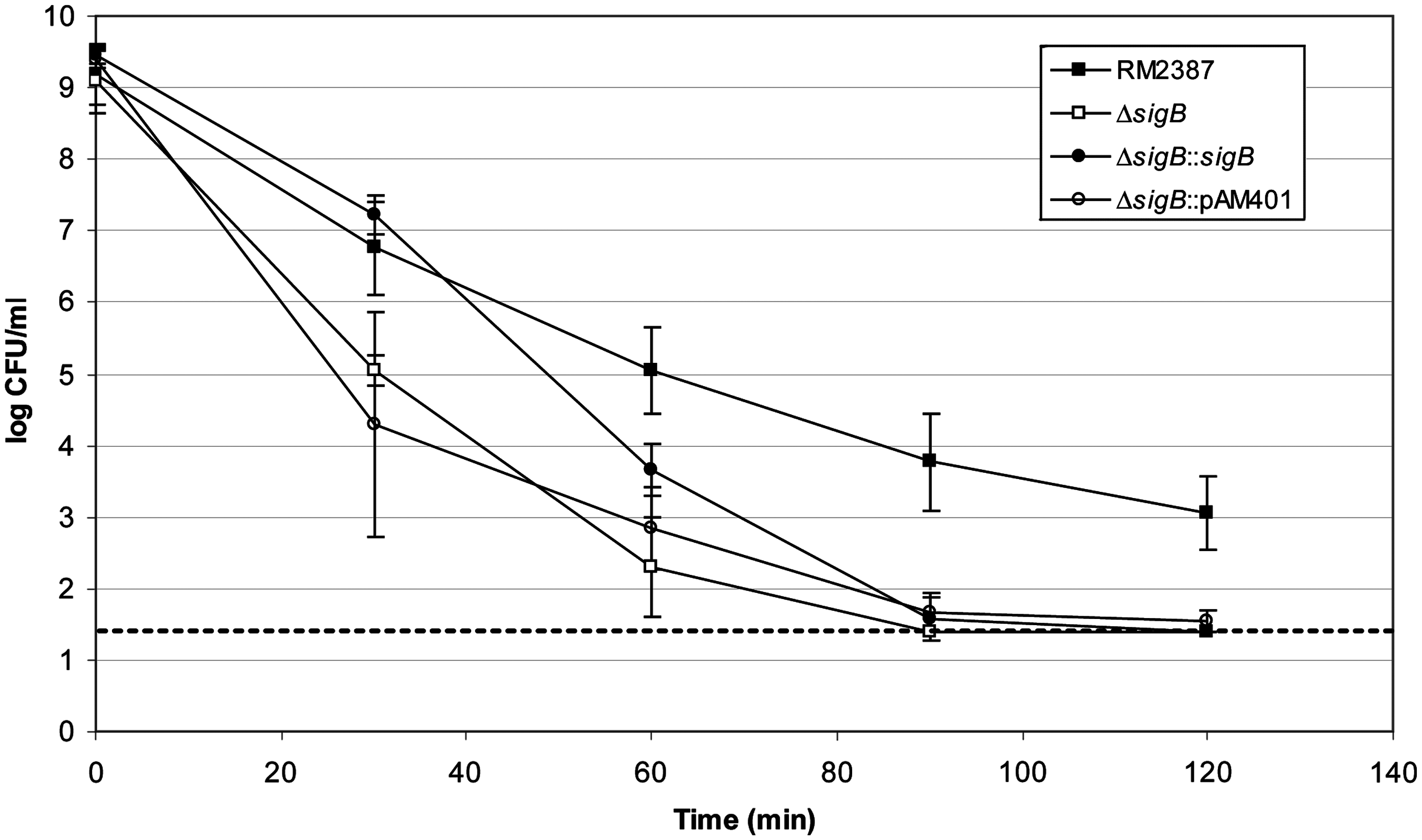
Acid resistance of Listeria monocytogenes strains. Shown are the log CFU/mL with standard deviation for each time point from representative experiments (n = 4). Dashed line shows limit of detection.
After oxidative stress, a survival rate of 84% was seen for the wild-type culture with survival of the mutant significantly decreased to 19% (p < 0.05). Addition of the wild-type sigB gene to the mutant increased survival to 51%, which was higher than the mutant with empty vector at 20% (p < 0.05).
Growth on radishes and in soil
Figure 2 shows a compilation of all experiments for the levels of the L. monocytogenes strains on mature radishes grown in contaminated soil. Regardless of the preparation method for the radishes, populations of the wild-type strain were up to 2 log CFU/g higher than the mutant (p < 0.001). Within strains there was no difference between radishes wiped clean of soil or washed under running water (p > 0.05). For the surface-sterilized radishes that were homogenized by stomaching, < 1 log CFU/g of the wild type was detectable; however, rolling these intact radishes over MOX plates yielded L. monocytogenes growth, indicating that there was incomplete surface sterilization rather than internalization. The mutant strain was never detected in homogenized radishes.
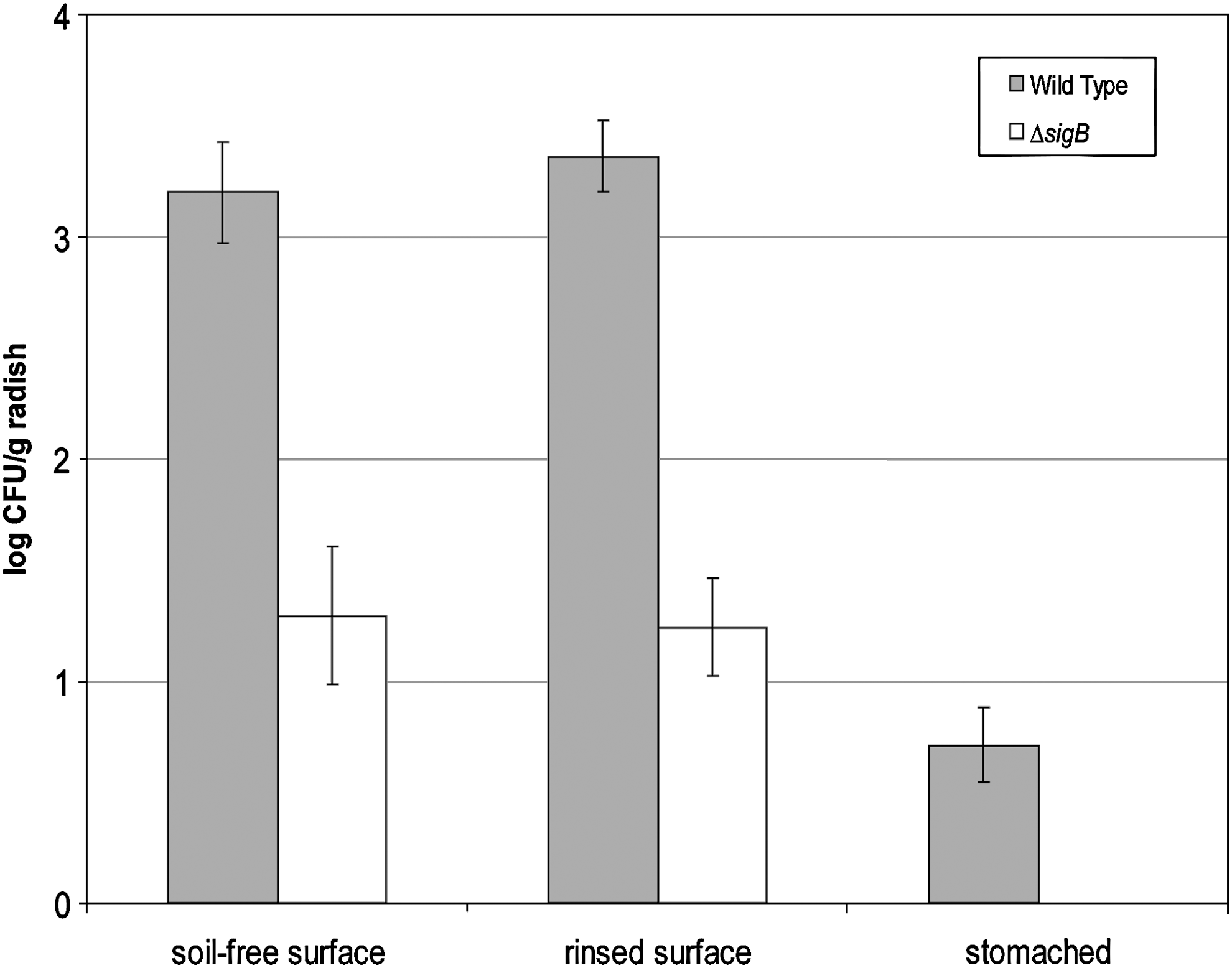
Levels of L. monocytogenes on radishes grown in contaminated soil. Shown are the results of compiled experiments with standard error.
The levels in soil were monitored during the 4-week trials (Fig. 3). By 24 h after inoculation, the mutant was 1 log CFU/g reduced from the wild type (p = 0.007), and it remained 1–2 log CFU/g lower throughout (p < 0.0001), despite being inoculated at the same levels. Both strains showed an increase through the first week in soil, with a higher rate for the wild type. The levels then reduced, and the slopes indicate a more severe decrease in the mutant. Interestingly, the strains had a 1–2 log difference both in soil and on mature radishes after 4 weeks (Fig. 2). Figure 4 shows the growth within the first 24 h after soil inoculation, and statistically there was no difference between the wild type and the mutant at 0 or 6 h (p > 0.05), but by 12 h there was a 0.5 log CFU/g soil difference (p < 0.001), which increased through 18 h (p < 0.05) to nearly a 1 log CFU/g difference by 24 h (p < 0.001). The complemented mutant was rescued partially. The complementation plasmid used for rescue is not stable. Without antibiotic selection 30%–40% of the population lost the plasmid (data not shown). By 24 h after soil inoculation the complemented mutant had intermediate levels between the wild type and the mutant. The transient nature of the complementation plasmid precluded its use over 4 weeks of growth with radishes, since antibiotics could not be added to the soil without affecting the growth of the plant, and the nature of the microbial community in the soil.
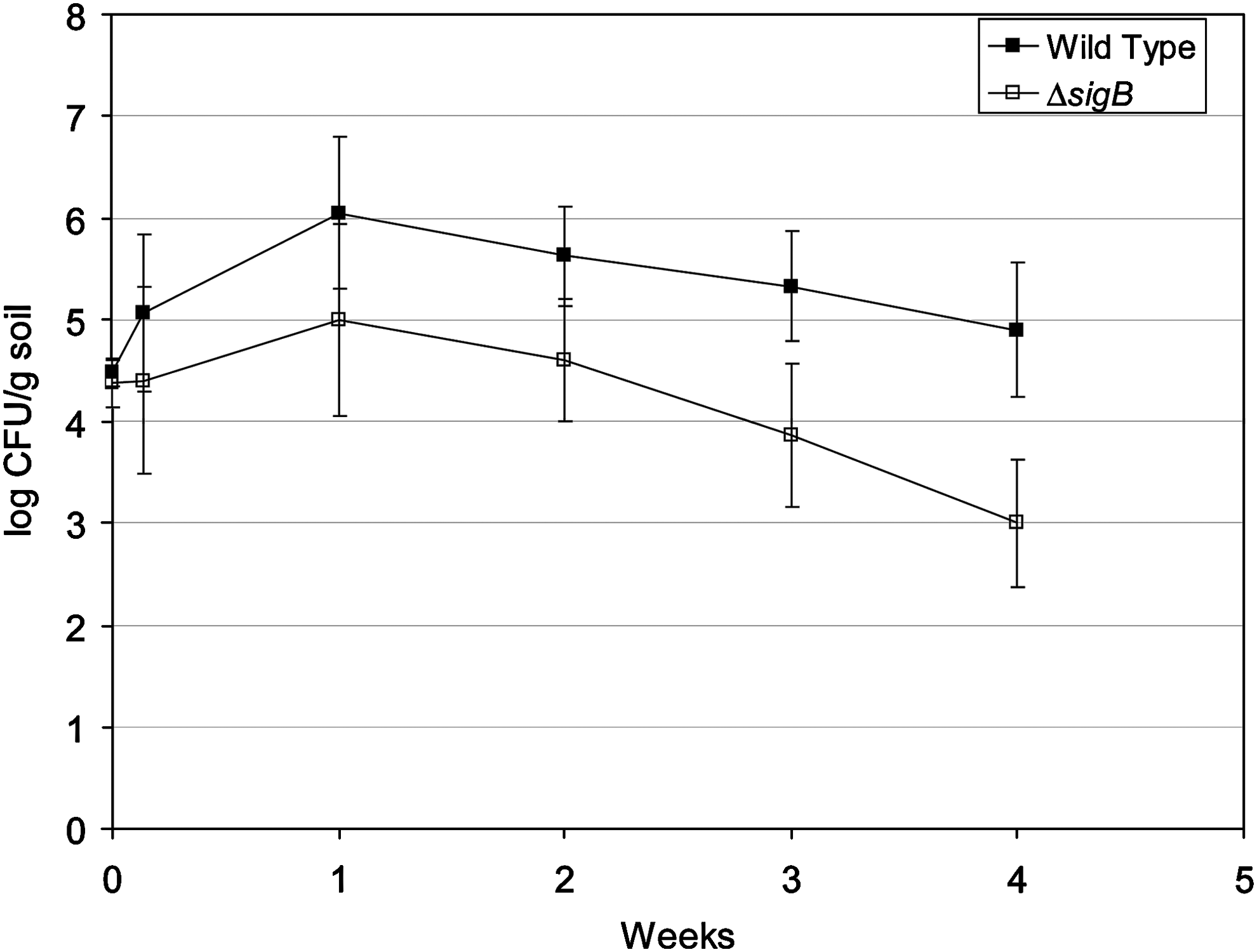
Levels of L. monocytogenes strains in contaminated soil over 4 weeks. Shown are the results of several experiments with standard deviations.
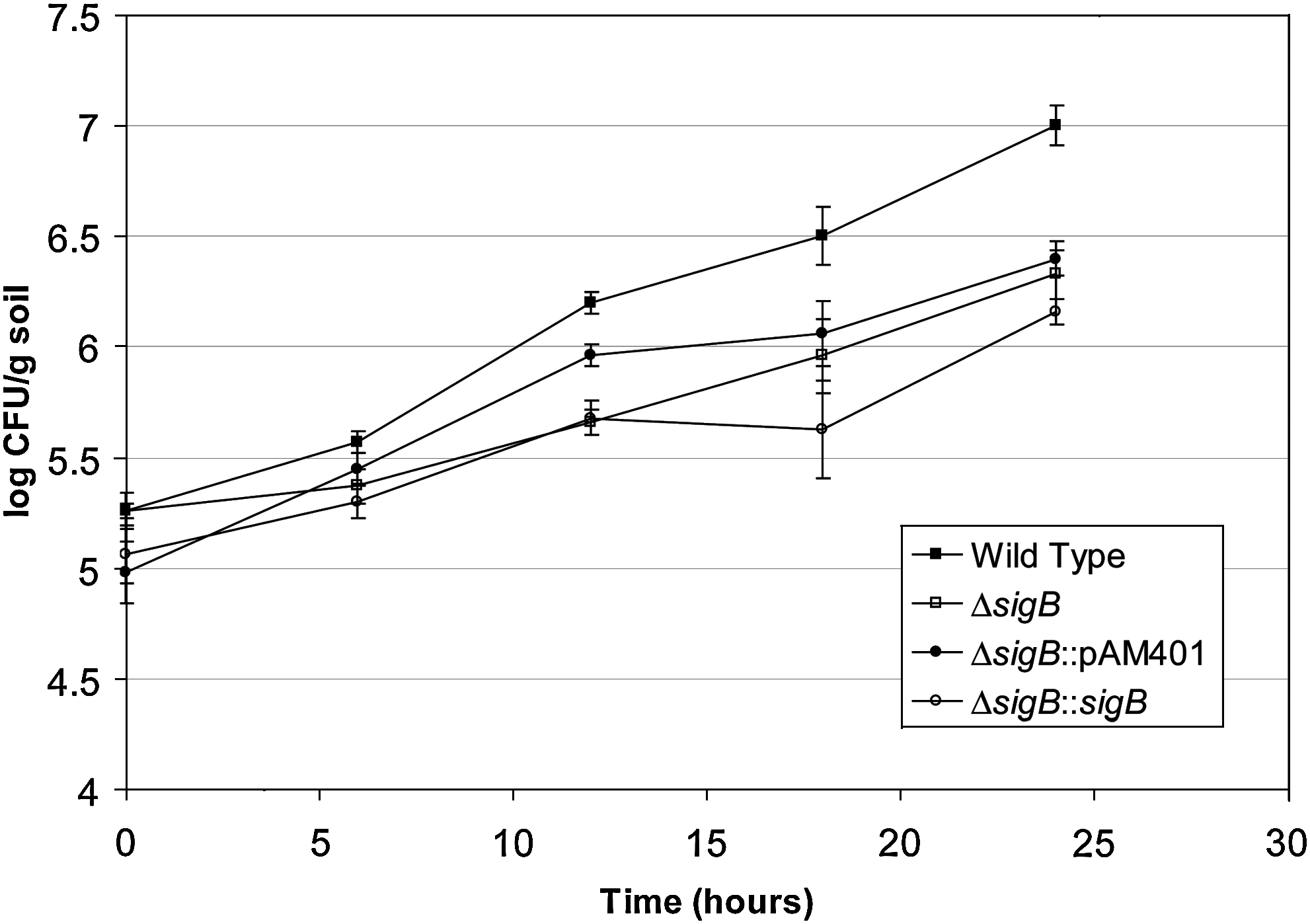
Levels of L. monocytogenes strains in soil over 24 h. Shown are the results of several experiments with standard error.
Discussion
Survival of an L. monocytogenes sigB operon mutant was assessed in soil and on radishes grown in contaminated soil in strain RM2387, an efficient plant colonizer (Gorski et al., 2008, 2009). The sigB mutant was an out-of-frame deletion; however, the downstream rsbX, involved in indirect negative regulation of sigB-dependent gene expression, was still transcribed (Shin et al., 2010). Even if RsbX functionality was affected, it would lead to defective negative regulation of sigB expression; therefore, it is likely that the phenotypes here were due to the absence of SigmaB. Due to the out-of-frame mutation, we conclude that the 7-gene sigB operon affects soil fitness.
The wild-type strain survived in the soil and on radishes at levels 1–2 log CFU/g higher than the mutant (Figs. 3 and 4). The deletion did not obliterate growth and survival in soil or on radishes, so non-SigmaB-mediated factors contribute to survival in this system. SigB controls the response to many, but not all stresses in L. monocytogenes, and it can vary between strains (Becker et al., 2000; Ferreira et al., 2001; Fraser et al., 2003; Kazmierczak et al., 2003; Moorhead and Dykes, 2003, 2004; Oliver et al., 2010).
After 24 h there was already a 1 log difference between the strains in soil, indicating an initial contribution by sigB to fitness (Figs. 3 and 4). The 1 log CFU/g difference was maintained until 3–4 weeks in soil when the mutant dropped another 1 log CFU/g, and after 4 weeks there was a 2 log CFU/g difference. Since this mathematical difference was reflected in levels on mature radishes it is likely that attachment and survival on the radish surfaces was not affected by the deletion. If sigB affected radish surface attachment and/or survival, then the difference between the wild type and the mutant on the radishes would be expected to be larger than the difference seen in the surrounding soil. Further, neither the wild-type nor the mutant could be wiped or washed off of the radish surface, so attachment and colonization factors were likely not SigmaB mediated. Schwab et al. (2005) showed that Sigma-B is not essential for attachment of L. monocytogenes to stainless steel.
Survival of bacteria in soil requires the ability to adapt to changing environments and stresses. DeAngelis et al. (2010) showed that a microbial population in soil was adapted to deal with fluctuating redox potentials imposed on the soil. Pseudomonas fluorescens needs an intact rpoS gene for full survival in the rhizosphere of bean plants (Stockwell and Loper, 2005), and P. putida needs it for root colonization (Miller et al., 2001). Roles for SigmaB for L. monocytogenes in soil were in the initial adaptation period (the first 24 h) and long-term survival. SigmaB is important for the transition into and survival in stationary phase. Another role for sigB for L. monocytogenes in this system could be related to growth phase, since sigB activity is induced in stationary phase (Ferreira et al., 2003; Oliver et al., 2010).
The transition to life inside the host is also mediated by sigma B, which plays a role in the transition from a saprophytic lifestyle to a virulent one in part by activating virulence genes (Gahan and Hill, 2005; Hain et al., 2008; Toledo-Arana et al., 2009). In addition to stress responses, sigB regulates genes involved in carbohydrate transport and metabolism, amino acid metabolism, ABC transporters, cell wall proteins, and virulence (Kazmierczak et al., 2003; Hain et al., 2008; Toledo-Arana et al., 2009). It is possible that sigB is active at the time of harvest but attempts to determine this with molecular analysis of plant-associated cells have proven difficult due to the strength of the attachment of the L. monocytogenes cells to the plant material. The strength of this attachment to a vegetable grown in contaminated soil should be noted for future experiments studying the efficiency of intervention methods to remove this pathogen from food surfaces.
Conclusions
The data indicated that the sigB operon has a function in adaptation and survival for L. monocytogenes in soil, but likely not in plant surface attachment. Other molecular factors not regulated by sigB play a significant role in the system as well. Additionally, the data indicate that the pathogen attaches very strongly to plant tissues when they are grown in contaminated soil, because it could not be washed off the radish surface.
Footnotes
Acknowledgments
We thank T. Chakraborty, Justus Liebig Universität Giessen, Giessen, Germany, for the gift of pAUL-A. We thank K. Nguyen and A. Tse for technical assistance. This research was funded by the U.S. Department of Agriculture (Agricultural Research Service CRIS project number 5325-42000-044-00D).
Disclosure Statement
No competing financial interests exist.