
Abstract
Free chlorine as hypochlorite is recommended to decontaminate fecally contaminated surfaces to control human norovirus (NoV). We evaluated the efficacy of sodium hypochlorite to decontaminate GII.4 NoV and three surrogates of human NoVs, feline calicivirus (FCV), murine norovirus (MNV), and coliphage MS2, on a fecally soiled stainless steel surface. Reduction of infectivity of FCV, MNV, and MS2 was measured by plaque assay and the decline of genomic copy numbers of GII.4 NoV by reverse transcriptase–polymerase chain reaction. Sodium hypochlorite solution at 5000 ppm could inactivate FCV by 3 log10 plaque forming units after approximately 1.9 minutes of contact time, but required longer exposure times of 3.2 and 4.5 minutes to reduce MNV and MS2 by 3 log10, respectively. However, detection of viral RNA by reverse transcriptase–polymerase chain reaction assay may not be reliable to estimate the effectiveness of sodium hypochlorite against human NoV. Of three NoV surrogates, FCV is not the most resistant of the virus tested for inactivation by hypochlorite and thus is not the worst-case model for estimating NoV inactivation. Although the use of 5000 ppm of hypochlorite for fecally soiled surfaces is effective, it may require longer exposure times of ≥3 minutes to control NoVs. Surface precleaning before hypochlorite disinfection is recommended to initially reduce the fecal organic load for better virus inactivation and should be a part of the environmental hygiene response measures during an NoV outbreak or where NoV fecal contamination of environmental surfaces is likely or suspected to be present.
Introduction
Human excretions such as stool and vomit are the main source of NoV environmental transmission, with viruses in feces having titers estimated to be up to 100 million virus particles per gram (Atmar et al., 2008). To interrupt transmission of NoV from contaminated surfaces, the use of chemical disinfectants is considered as a key intervention measure (Barker et al., 2004). However, the optimum conditions for surface disinfection of NoV have not been adequately defined and remain poorly documented.
Due to the absence of a cell culture method to measure human NoV infectivity, the use of cultivable surrogate viruses such as feline calicivirus (FCV), murine norovirus (MNV), and coliphage MS2 have been used. In addition, reverse transcriptase–polymerase chain reaction (RT-PCR) assay methods to measure decline of viral RNA to evaluate the effectiveness of commercial disinfectants against human NoV contamination on environmental surfaces have gained considerable interest and use (Doultree et al., 1999; Barker et al., 2004; Duizer et al., 2004; Park et al., 2007). Different virus assays and surrogate viruses have contributed to the understanding of the likely inactivation responses of human NoV exposed to disinfectants on environmental surfaces and in water and foods. However, the interpretations and comparisons of these data are very difficult, because of the lack of standardization in virus exposure and assay methods and the lack of comparative studies simultaneously including human NoV and different candidate surrogate viruses
In this study, the effect of hypochlorite on the infectivity of three non-enveloped NoV surrogate viruses, FCV, MNV, and MS2, as well as human GII.4 NoV was tested after drying onto a stainless steel coupon as a model of nonporous fomitic surface. As human NoV cannot be cultured, RT-PCR was used to investigate the effect of hypochlorite on all of these viruses.
Materials and Methods
Viruses
The human GII.4 NoV-positive stool sample was obtained from an outbreak at the University of North Carolina campus in 2004, which were aliquotted and frozen. This frozen material was freeze-thawed no more than twice in order to maintain virus infectivity.
Bacteriophage MS2
Bacteriophage MS2 (ATCC No. 15597-B1) was cultivated using Escherichia coli (E. coli) Famp (ATCC No. 700891) with minor modifications from the original protocol (USEPA, 2001b). In brief, log-phase E. coli cells grown in Lauria-Bertani (LB) broth at 37°C were infected with MS2 at a multiplicity of infection (MOI=ratio of infectious virus particles to cells) of 0.05 to 0.5 and incubated overnight at 37°C. MS2 was partially purified from infected cell lysates by the recovery of the resulting supernatant after moderate-speed centrifugation (3000 g, 20-minutes, 4°C). The titer of the partially purified MS2 stock was approximately 1010 plaque forming units (PFU) per milliliter.
Mammalian viruses, host cells, and plaque assays
MNV (strain CW3) was kindly provided by Dr. Skip Virgin, Washington University School of Medicine (St Louis, MO), and FCV was strain F9 ATCC No.VR-782. MNV was propagated and assayed in RAW 264.7 cells (ATCC No. TIB-71, Manassas, VA) and FCV was propagated and assayed in Crandell Reese Feline Kidney cells (CRfK ATCC No. CCL-94), as previously described (Park et al., 2011). The infectivity titers of MNV and FCV stocks were approximately108.0 and 108.9 PFU/mL, respectively. Virus stocks were stored in aliquots at −80°C and each aliquot was thawed for use only once.
RT-PCR assays for MNV, FCV, MS2, and GII.4 NoV
Viral RNA was extracted from sample suspensions using the QIAamp viral mini kit, according to the manufacture's instructions (QIAGEN, Inc, Valencia, CA). Viral RNA (MNV, MS2, and GII.4 NoV) was detected by RT-PCR assay using the Qiagen One-step RT-PCR kit (QIAGEN, Inc), as previously described (Park et al., 2007). For FCV strain F9, forward primer 5′- TGCTTTCCACAAACTCAACC-3′ and reverse primer 5′-CCCAGATCATCCTTCTTTTCC-3′, corresponding to nucleotides [nt] 4069 to 4298, were used to amplify viral RNA. RT-PCR amplification conditions for FCV were 45°C for 30 minutes and 94°C for 15 minutes, followed by 40 PCR amplification cycles consisting of 30 seconds at 94°C, 1 minute at 50°C, and 30 seconds at 72°C, and a final extension step at 72°C for 10 minutes.
To quantify the RNA titer of each virus, serial 10-fold dilutions of RNA were assayed by RT-PCR to determine the highest dilution of virus stock that is positive, as a dilution end point titer. The average titer and standard deviation of four replicates was computed using the Thomas approximation and the DeMan procedure, respectively (Peeler et al., 1992). Amplification reaction products (15 μL) were resolved by electrophoresis (1 hour, 120 V) on 2% agarose gels, stained with ethidium bromide, and observed by ultraviolet transillumination.
Carrier test
No. 4 finish polished steel coupons of size of 1×1 cm2 were used for the carrier test to measure the efficacy of sodium hypochlorite on three surrogate viruses and human GII.4 NoV (Park et al., 2007). A 10% (w/v) stool suspension consisted of 100 mg/mL solid human stool having 106.6 RNA titer/mL of human GII.4 NoV, and three NoV surrogate viruses, MS2, FCV, and MNV, at the titers of 108.3 (109.4), 106.7(107.0), and 107.4 (108.8) PFU/mL (RNA titer/mL), respectively. Commercial Clorox sodium hypochlorite bleach (6.2% [w/v]) was diluted in deionized water to make a stock solution at 5000 ppm free chlorine concentration and pH 7.0. Free chlorine concentration was measured by the DPD colorimetric method using a portable chlorine colorimeter (Hanna Instrument, Woodsocket, RI). Prepared stock solution was further diluted in deionized water to make hypochlorite solutions at free chlorine concentrations of 250, 500, 1000, and 2500 ppm, respectively. Before the carrier test, effectiveness of sodium thiosulfate to neutralize biocidal activity and cytotoxicity of sodium hypochlorite solution on each cell line were validated (Park et al., 2010). For the carrier tests, aliquots of 25 μL of a virus pool containing all four viruses were applied onto the center of each of three stainless steel coupons and extensively air dried (1–1.5 hours), as previously described (Park et al., 2007). The carriers were then transferred to individual wells of a 24-well plate, each well having 1.2 mL of pH-adjusted hypochlorite solution (pH 7.0) (or phosphate-buffered saline as a negative control) at various target concentrations. The dose–response relationship and inactivation kinetics of each virus by sodium hypochlorite concentrations were determined by exposing dried viruses on stainless coupons to sodium hypochlorite concentrations ranging from 250 to 5000 ppm for 2 minutes or to sodium hypochlorite solutions at 500 and 5000 ppm for up to 4 and 10 minutes, respectively. Following specific contact times, 0.275 mL of 16% beef extract (pH 8.0) and 0.025 mL of 30% sodium thiosulfate were added to simultaneously to neutralize the hypochlorite and facilitate elution of virus from carrier. The plates were mechanically rotated for 15 minutes, and coupons were mechanically washed by repeated pipetting for 10 seconds, after which the solution was transferred into sterile 1.5 mL microcentrifuge tubes and either analyzed directly by infectivity assay (Park, et al., 2011) or by RT-PCR assay or were kept at −80°C until assayed.
Data analysis
The virucidal activity of the hypochlorite dilutions was determined by calculating the log10 virus reduction as Nd/N0, where N0 is the titer of infectious viruses (or RNA titer) of the untreated sample and Nd is the titer of infectious viruses (or RNA titer) of the chlorine-treated sample. A linear regression was also used to determine the inactivation kinetics as a function of time, the differences in inactivation rates between viruses for different test conditions, or the relationship between reduction in infectivity titer and chlorine dose using SAS software, as previously described (SAS 9.2; SAS institute Inc, Cary, NC) (Peeler et al., 1992; Moore and McCabe, 1998; Park et al., 2011).
Results
In this study, we measured the inactivation magnitudes of MNV, FCV, and MS2 on a fecally soiled solid stainless steel surface treated with sodium hypochlorite solution. We tested different concentrations (250, 500, 1000, 2500, and 5000 ppm) of sodium hypochlorite. The reduction of infectivity titer was plotted as a function of applied chlorine dose. The titer of MNV and FCV decreased proportionally with an increased concentration of hypochlorite with slope rates (log10 PFU/mL per mg/mL [free chlorine concentration]) of −0.0003 (r 2=0.966) and −0.0006 (r 2=0.985), respectively (Fig. 1). In contrast, the titer of MS2 decreased by 0.6 log10 PFU/mL after exposure to 250 ppm of chlorine. However, for free chlorine doses ranging from 250 to 5000 ppm, the MS2 titer slowly decreased with declining rates (log10 PFU/mL per mg/L [free chlorine concentration]) of −0.0002 (r 2=0.977). The reduction of the titer of the three viruses was lower than 1.3 log10 PFU/mL, if applied chlorine concentration was ≤2500 ppm. However, exposure to 5000 ppm of sodium hypochlorite solution resulted in 3.2 log10 inactivation for FCV, whereas the MS2 and MNV titers decreased by 1.6 and 1.5 log10, respectively.
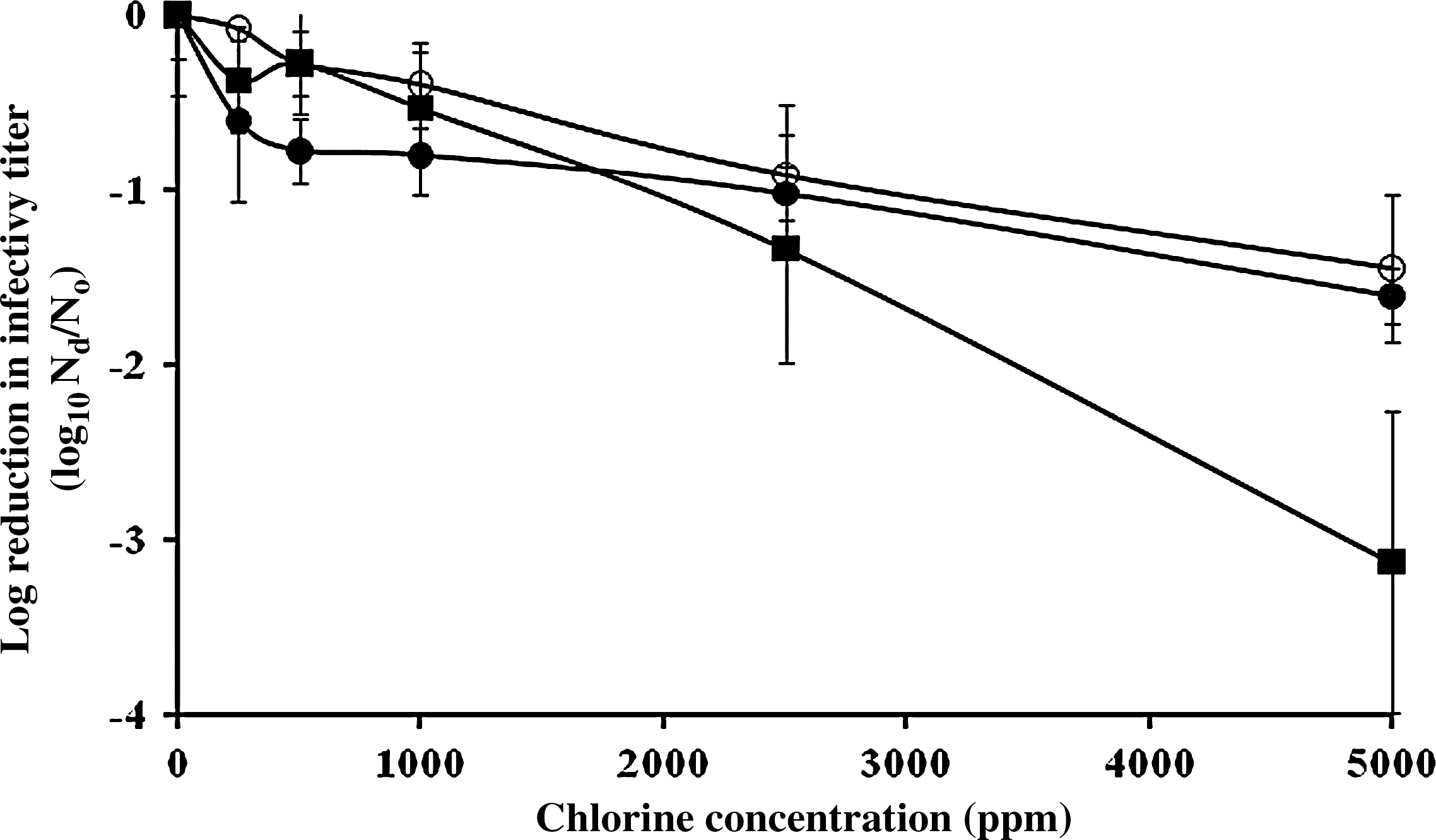
Dose–response relationship of NoV surrogate viruses in 10% stool suspension by sodium hypochlorite at free chlorine doses ranging from 250 to 5000 ppm. Each symbol is designated as follows: MS2 (•), MNV (○), and FCV (▪). Exposure time is 2 minutes and infectivity data are the mean of six replicates from two independent experiments (three replicates per experiment). Bars on the graphs indicate standard deviations. NoV, norovirus; MNV, Murine norovirus; FCV, Feline calicivirus.
The inactivation rates of FCV, MNV, and MS2 by 500 and 5000 ppm sodium hypochlorite on a fecally soiled stainless steel carrier were determined, as shown in Figures 2 and 3, respectively. The titer of infectious MNV, MS2, and FCV after 10 minutes of treatment with 500 ppm of sodium hypochlorite on fecally soiled stainless steel carrier was reduced by 0.9, 0.8, and 1.9 log10 PFU/mL, respectively (Fig. 2a). Based on regression analysis, the infectivity reduction rates of MNV and MS2 were not statistically different from each other (p=0.8703), but both were significantly lower than that of FCV with p<0.05. After 10 minutes of exposure to 500 ppm sodium hypochlorite, the detectable RT-PCR titers of MS2 remained unchanged, whereas the titers of MNV, FCV, and GII.4 NoV were reduced by 0.2, 0.8, and 0.6 log10 RNA copy number/mL, respectively (Fig. 2b).
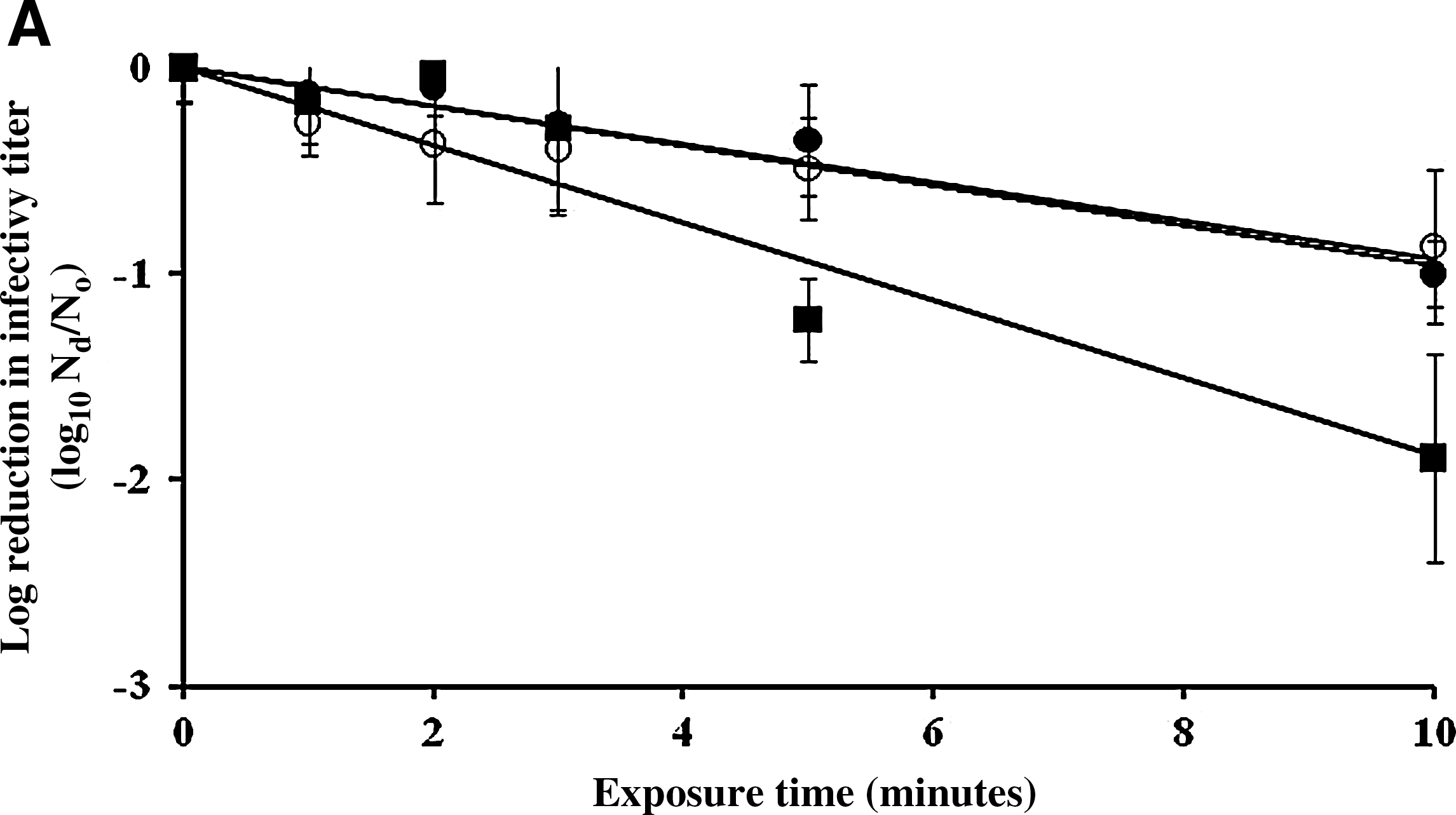
Surface inactivation kinetics [
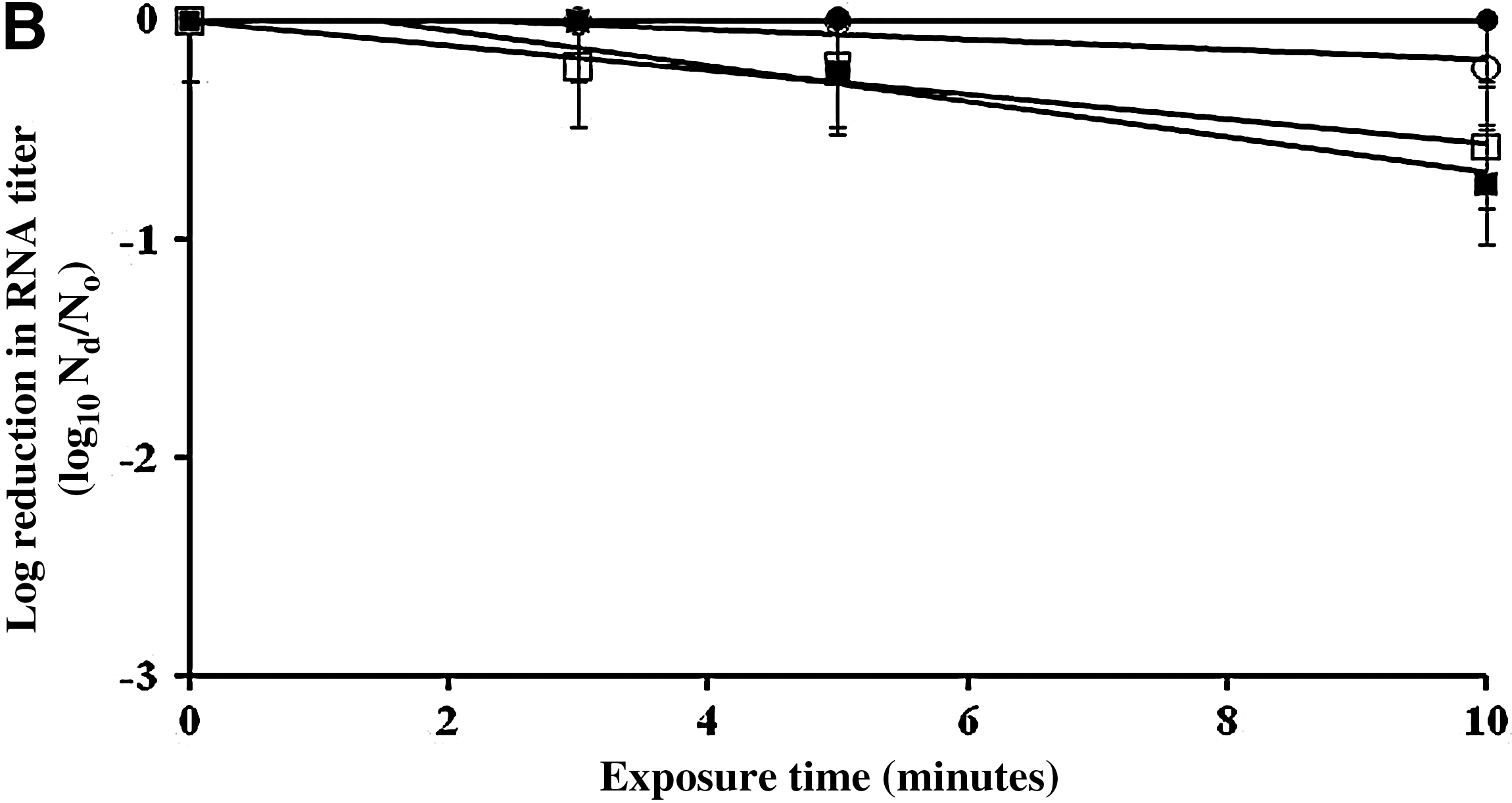
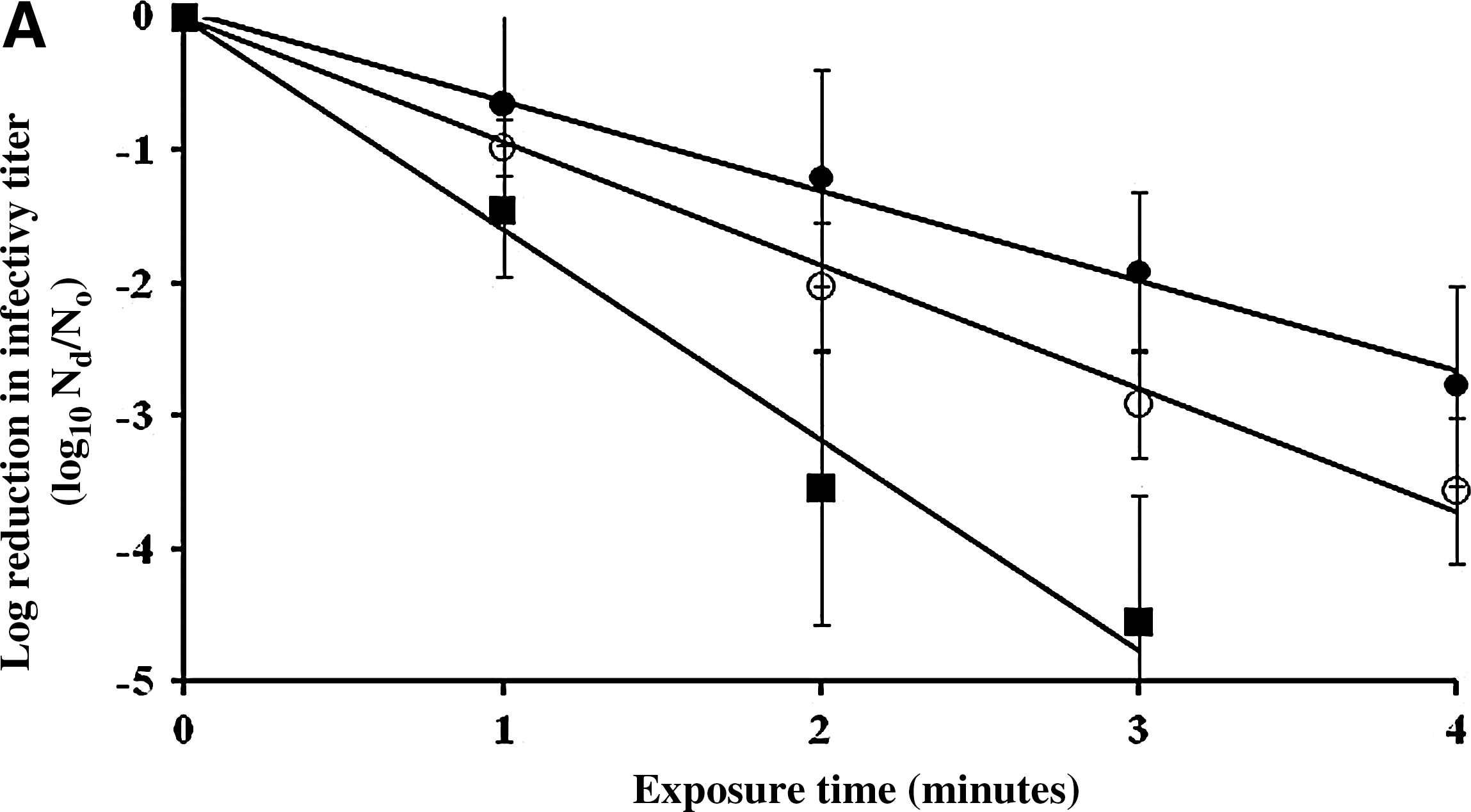
Surface inactivation kinetics [
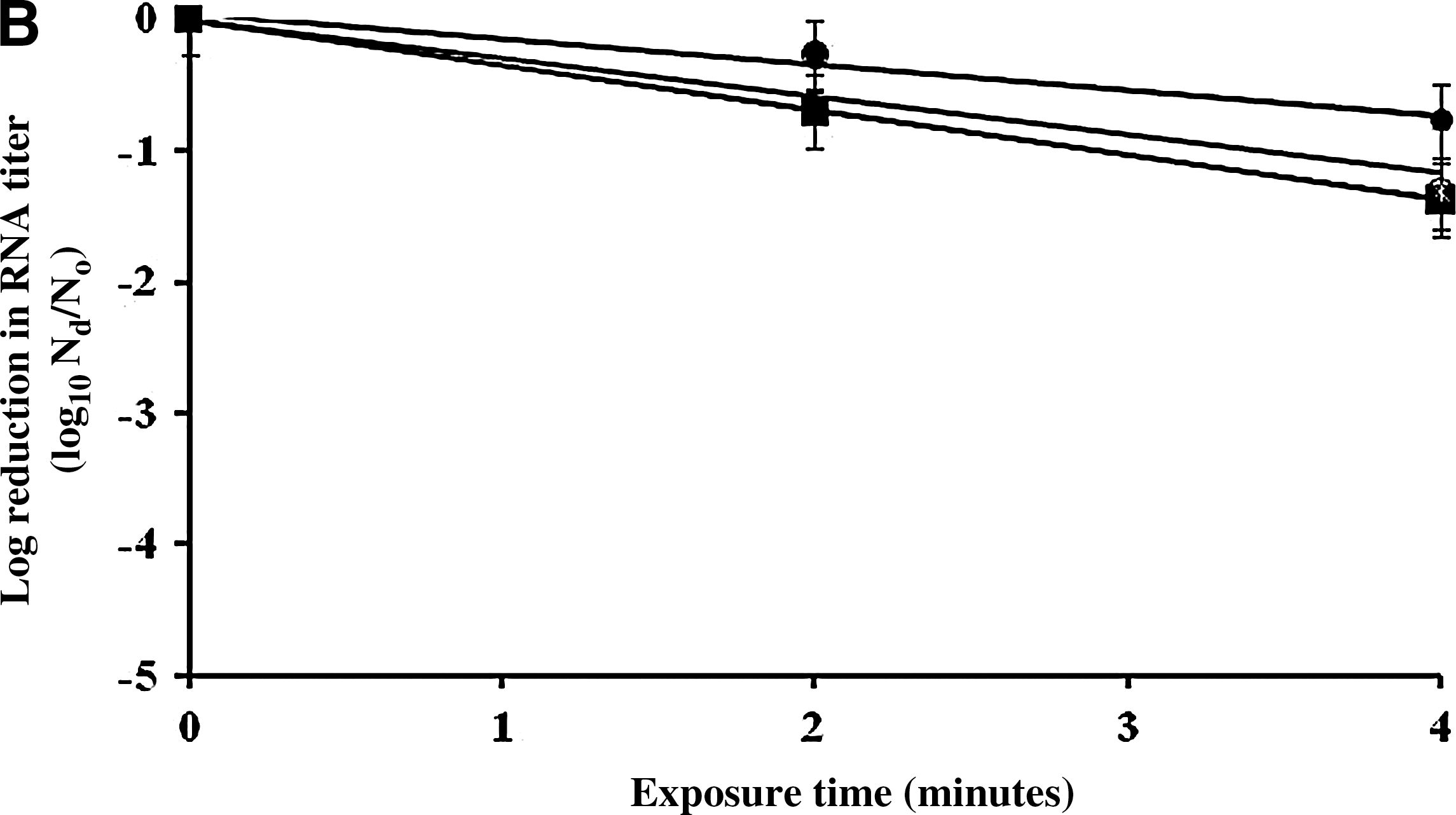
At 5000 ppm of sodium hypochlorite, the inactivation rates for all viruses were greater than those for 500 ppm sodium hypochlorite, and they were significantly different from each other with p<0.05. FCV was rapidly inactivated by 3.2 log10 PFU/mL after 2 minutes and by 4.5 log10 after 3 minutes (Fig. 3a). However, 3.2 minutes or longer exposure (4.5 minutes) were needed to inactivate MNV and MS2 by more than 3 log10 PFU/mL. The infectivity loss of FCV after 3 minutes of exposure to 5000 ppm of sodium hypochlorite was significantly greater than that of both MNV and MS2 (p=0.0347 and 0.00548), respectively. RNA titers of MS2, FCV, MNV, and GII.4 NoV were reduced by 0.8, 1.4, 1.3, and 1.4 log10 RNA copy number/mL, respectively, after 4 minutes of exposure (Fig. 3b). However, there were no statistically significant differences in RNA titer reductions between viruses (p>0.05).
Discussion
Sodium hypochlorite is one of the most widely used chemicals to disinfect contaminated surfaces for a broad range of microbes, including viruses, bacteria, and protozoa (Rutala and Weber, 1997). Importantly, virucidal activities against human NoV on surfaces have not been adequately documented (Doultree et al., 1999; Barker et al., 2004; Duizer et al., 2004). In fact, most of these studies were not consistent in infectivity assays (e.g., surrogate viruses or RT-PCR), which makes it difficult to compare among studies or relate their finding to human NoV. Moreover, the result of these previous studies might not predict the virucidal activity of hypochlorite against NoV on fecally soiled surfaces because their test conditions of lower organic loads, longer exposure times, and the use of either aqueous virus suspension or partially dried virus suspensions were unlikely to simulate field conditions (Sattar et al., 2003).
The assessment of the virucidal activity of chemical disinfectants is generally obtained under defined test conditions, which may be different from conditions encountered in the field. For example, contact times used in most laboratory studies are typically in the range of 1–10 minutes, which is much longer than in a field situation where the contact time between a virus contaminated area and a disinfectant is estimated to be between 1 to 3 minutes or even much less (Sattar et al., 2003). To achieve at least a 3 log10 infectivity reduction for FCV, MNV, and MS2 by 5000 ppm hypochlorite, exposure times of approximately 1.9, 3.2, and 4.5 minutes were needed, respectively. Although the recommended 1–3 minutes of exposure to 5000 ppm hypochlorite-inactivated FCV by 3 log10, shorter exposure times of one minute or less are unlikely to inactivate any of the surrogate viruses by 3 log10. Given its greater susceptibility to hypochlorite inactivation compared to MS2 and MNV, FCV does not represent the most resistant surrogate virus and thus its use may overestimate the efficiency of hypochlorite to decontaminate NoV infectivity on surfaces.
The log10 decrease of RT-PCR amplifiable RNA titers that correspond to a 3-log10 reduction of virus infectivity after exposure to hypochlorite was estimated to be approximately 0.8, 0.7, and 0.9 log10 RNA copy numbers/mL for MS2, FCV, and MNV, respectively (Fig. 2b). Because these values seem reasonably consistent between the different viruses, they seem to provide an indirect estimate of the NoV reduction. However, caution in predicting infectivity titer reduction by this approach is recommended, because RNA titer reduction corresponded to a wide range of infectivity titer reductions, which also varied by hypochlorite doses and virus strains. As in previous studies, RT-PCR assay was not reliable to predict NoV infectivity or the magnitude of its reduction in infectivity titer by disinfection, by consistently underestimating virus infectivity reductions achieved by disinfection treatments (Sobsey et al., 1998; Park et al., 2007, 2010).
In this carrier test as previously described, a high volume ratio (1:48) of sample volume to disinfectant volume was employed (Abad et al., 1997; Park et al., 2007). Based on interpolation or extrapolation of the regression curves for inactivation kinetics of the three viruses at two concentrations of hypochlorite (500 and 5000 ppm), Ct value ranges [disinfectant concentration (mg/L)×contact time (minutes) value ranges, as mg/L-minute] to achieve 3 log10 reductions for each virus were estimated to be 1.56×104 (500 ppm) to 1.60×104 (5000 ppm), 1.63×104 (500 ppm) to 2.26×104 (5000 ppm), and 0.79×104 (500 ppm) to 0.94×104 (5000 ppm) for MNV, MS2, and FCV, respectively. Hence, the Ct values were essentially similar, irrespective of being based on the lower or higher hypochlorite concentration tested. There was not a dramatic decline of virus infectivity on the test surfaces after up to 4 minutes of exposure to 5000 ppm hypochlorite and up to 10 minutes of exposure to 500 ppm hypochlorite. All tested viruses dried onto carriers appeared to be more resistant to sodium hypochlorite than aqueous suspended viruses, as estimated Ct values (mg/L-minute) of aqueous suspended viruses by hypochlorite were much less than 166 mg/L-minute (data not shown) (Shin et al., 2008).
NoVs are shed primarily in an organic matrix such as feces (Atmar et al., 2008). This matrix may influence the disinfection efficacy of hypochlorite by protecting viruses from exposure to hypochlorite or by chemically interacting with and neutralizing it (Rutala and Weber, 1997). Previously, a 1% dilution of a NoV stool was used to test the virucidal activity of free chlorine as hypochlorous acid against MS2 on stainless steel coupons (Park et al., 2007). In that study, Ct value to achieve 3-log10 reduction of MS2 dried on carrier surfaces was estimated to be approximately 94, more than 173-fold lower than the Ct values obtained in this study (Park et al., 2007). However, in both studies, a >4-log reduction of aqueous suspended MS2 in 1% and 10% stool suspension was obtained in <20 seconds of exposure to free chlorine at 18.8 and 500 ppm, respectively. Both studies strongly demonstrate that the resistance of MS2 to free chlorine is highly influence by fecal matters present. More importantly, the protective effect of higher fecal matter concentration was higher for MS2 dried on the surface than in aqueous suspension for free chlorine disinfection. However, the difference in the resistance of MS2 to free chlorine in both studies cannot be fully explained solely by fecal matter concentration as an organic load. Variability in the composition of feces used in these studies might be another factor (Sattar et al., 2003). Additionally, it is possible that the larger mass of particulate fecal matter in the test suspension could allow viruses to remain attached to the carrier surface more firmly and for a longer period, making them less susceptible to inactivation by free chlorine.
Currently, U.S. Environmental Protection Agency (USEPA) recommends only FCV strain as an NoV surrogate to evaluate commercial disinfectants for label claims of antimicrobial activity against human NoV (USEPA, 2001a). However, numerous reports have demonstrated that FCV has a different inactivation pattern than other surrogate viruses for human NoV for disinfectants (Cannon et al., 2006; Park et al., 2010, 2011). In addition, our data demonstrate that FCV is not the most resistant surrogate virus to measure inactivation on surfaces and hence does not represent the worst-case model to estimate the level of inactivation of human NoV by hypochlorite. Therefore, the use of multiple surrogate viruses rather than a single surrogate virus is recommended in order to obtain more reliable information on the effectiveness of disinfectants on human NoV.
To our knowledge, this is the first report on measuring the effectiveness of sodium hypochlorite against three surrogate viruses on fecally soiled surfaces. Additionally, our study strongly suggests that more caution should be taken to disinfect NoV on fecally soiled surfaces. The use of 5000 ppm of hypochlorite for disinfection of fecally soiled surfaces may be a reasonable choice, but may still require longer exposure times of ≥3 minutes to extensively inactivate NoVs by 3 log10, regarded as a satisfactory reduction level for viral surface contamination (Springthorpe et al., 1986). Thus, surface pre-cleaning before hypochlorite disinfection is highly recommended to reduce the fecal organic load and should be a part of the environmental hygiene response measures during a NoV outbreak or where NoV fecal contamination of environmental surfaces is likely or suspected to be present.
Footnotes
Acknowledgments
We are grateful for the technical assistance of Douglas Wait and Dorothy Thompson of University of North Carolina (UNC) and thank Jan Vinjé (CDC) for carefully reviewing the article.
Disclosure Statement
No competing financial interests exist.