
Abstract
All hosts, including humans, can be infected by any one of the three forms of the parasite Toxoplasma gondii that correspond to three morphological stages: tachyzoite, bradyzoite, and sporozoite form. Felids are definitive hosts for T. gondii, which is an intracellular pathogen that infects a wide range of warm-blooded intermediate hosts. Toxoplasmosis is a disease where the interest of the diverse medical and veterinary specialties converge. Awareness needs to be increased that toxoplasmosis can induce clinical disease not only in immunocompromised patients or through congenital infections, but also in healthy patients. This is a review article that aims at illustrating why toxoplasmosis should be regarded a veterinary public health issue and how veterinary practitioners can contribute in controlling the infection.
Introduction
Etiology and Pathogenesis
T. gondii—from the Greek, to'xon: bow and plasma: shape, whereas gundi, Ctenodactylus gundi, is the North African rodent from which Toxoplasma was first isolated in 1908 (Weiss and Kami, 2007)—is a cosmopolitan protozoon (Anonymous, 2009b) with no host specificity in the asexual stage (it can parasitize all mammals, including humans and felids), whereas in the sexual stage it is specific to felids where it becomes localized in the intestine. Toxoplasma can become systemic via the blood stream and localize in vital organs, muscle tissue, and the nervous system. All hosts, including humans, can be infected by any one of the three forms of the parasite that correspond to three morphological stages: tachyzoites (in pseudocysts and free in exudates, blood, secretions, and excretions), bradyzoites (in tissue and organ cysts) (Fig. 1), or sporozoites (in oocysts in cat feces) (Dubey, 2008; Innes, 2010).
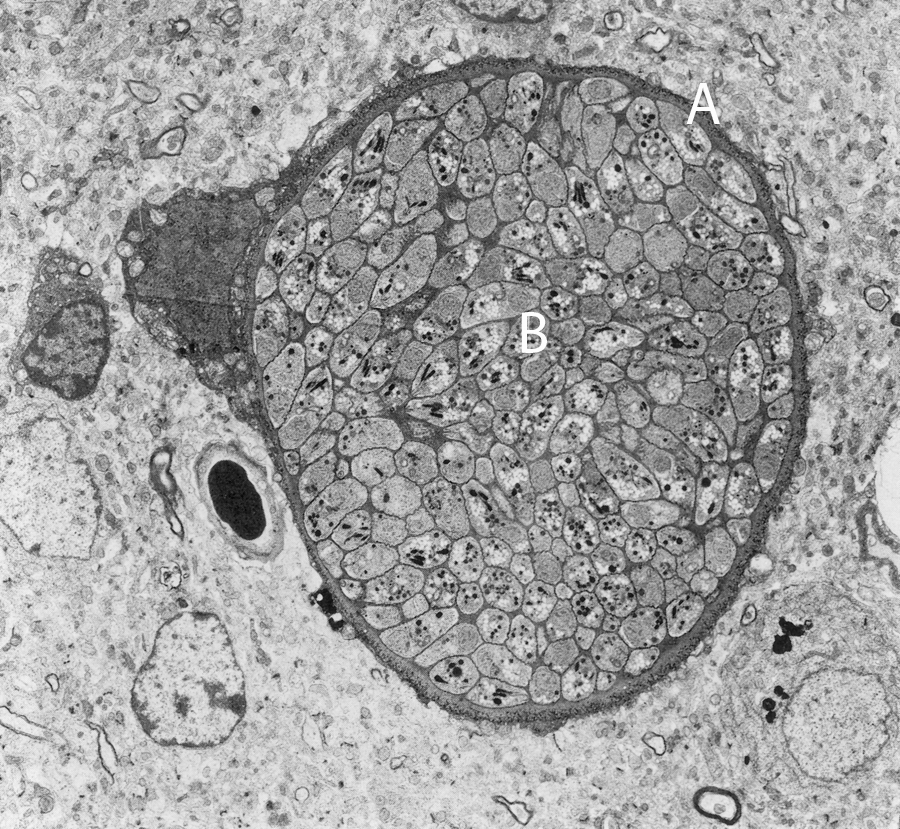
Tissue cyst in mouse brain on electronic transmission microscopy (Baldelli, 1983). A, cyst wall; B, bradyzoites. Kindly supplied by Dr. Marcello Tosti, Veterinarian, Perugia.
The sexual stage takes place in the intestinal epithelium of a felid (Fig. 2). The epithelial cells are infected with tachyzoites or bradyzoites present in ingested infected tissue or by sporozoites (in oocysts in cat feces) eliminated in another felid's feces that have contaminated food (or water) or by direct transmission of oocysts. Schizogony (endopolygeny) takes place in the intestinal mucosa and schizonts form in the intestinal tract, in 3 to 15 days, to give rise to gametes. After fertilization, the female gamete becomes a zygote and then an immature oocyst. Oocysts are first expelled in the intestinal lumen and then to the outside in feces: sporogony and maturation of the oocysts can take place, in 1–5 days, only in the outside environment (Lappin, 2010).
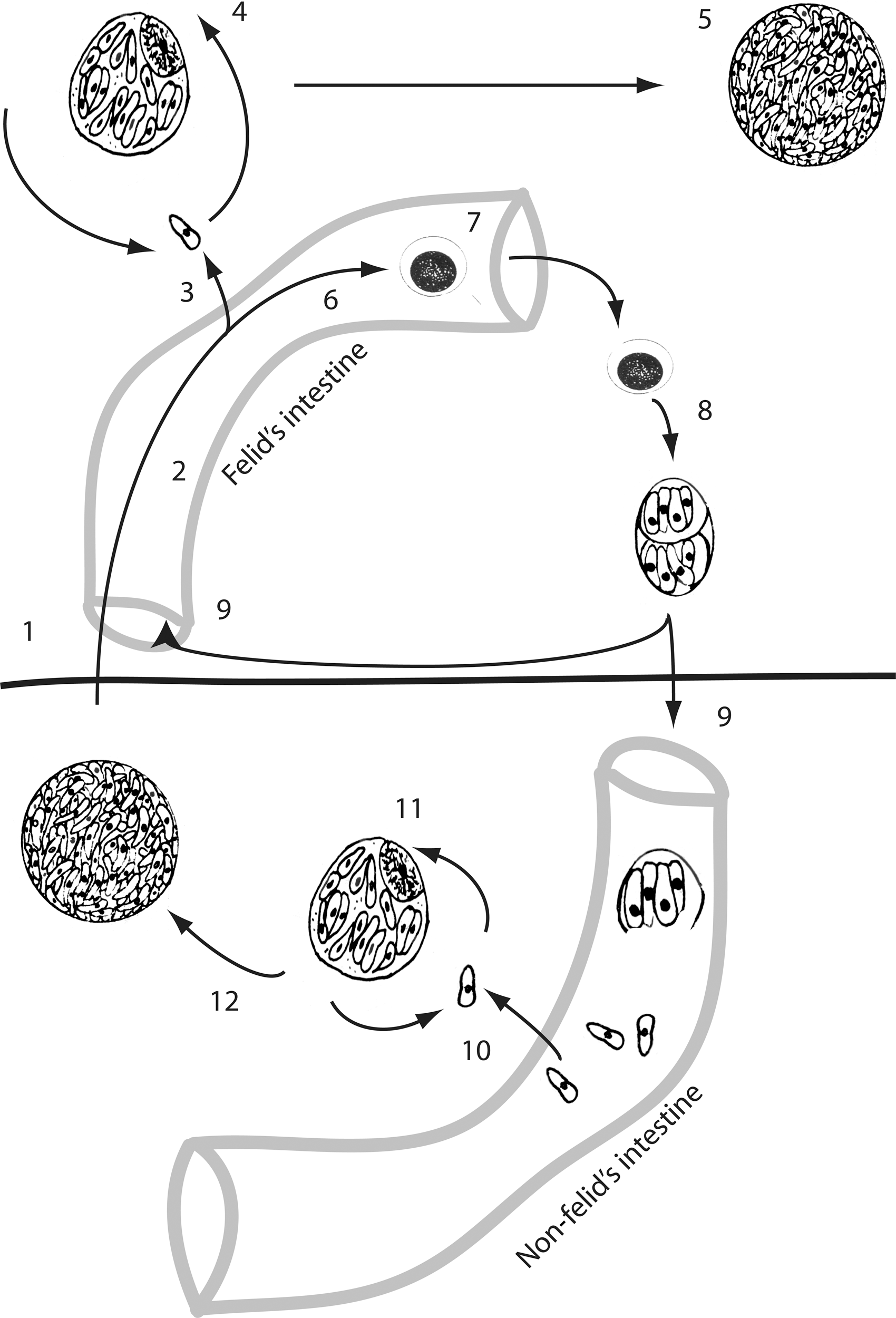
Life cycle of Toxoplasma gondii. Development in the intermediate host is illustrated below the horizontal bar; development in the definitive host is illustrated above the horizontal bar: (1) cyst ingested by a felid, (2) schizogony in the felid's intestinal mucosa, (3) diffusion of zoites to the felid's body, (4) pseudocyst, (5) cyst, (6) gamogony, (7) oocyst formation and expulsion, (8) sporogony and maturation of the oocysts that contains two sporocysts, each containing four sporozoites, (9) oocyst ingested by hosts, (10) diffusion of zoites to the non felid's body, (11) pseudocyst, and (12) cyst.
In intermediate hosts, T. gondii undergoes two phases of asexual development: in the first phase, tachyzoites multiply rapidly by repeated endodyogeny in many different types of host cells; in the second phase, tachyzoites of the last generation determine the formation of tissue cysts where bradyzoites multiply slowly by endodyogeny. Tissue cysts are the terminal life-cycle stage in the intermediate host and are immediately infectious (Dubey, 2008).
Tissue cysts can also be found in felids when some zoites (sporozoites, tachyzoites, or bradyzoites) break-up the intestinal lamina propria, get phagocytized, and multiply by endodyogeny (Anonymous, 2007).
Cats
Seroprevalence varies according to age and lifestyle, and higher values are found in strays than in house cats (Dubey et al., 2008a). In Europe data made available in 2009 show results of a survey with only limited indications on how the analyses were done (Anonymous, 2009a). In the United States a prevalence of 31% in >12,000 cats that underwent serological tests has been reported (Vollaire et al., 2005).
Since the large amount of figures available to the scientific community does not come from coordinated international or national studies, a filter suited to collect data on clinical toxoplasmosis in cats could be veterinary clinics that might also play an important role in controlling this disease. It must be said that in the absence of evident symptoms, the diagnosis is never easy, even in cases where the veterinarian has included toxoplasmosis in the possible differential diagnoses, especially when there is no possible recourse to laboratory confirmatory testing, which can be very expensive and not always available in many veterinary clinics (Lappin, 2000; McConnell et al., 2007; Falzone et al., 2008).
Most infected cats are asymptomatic, whereas clinical toxoplasmosis is mostly manifested in pneumonia, and in cats that subsequently died the most common signs were sensory depression and anorexia (Dubey, 2008). Other consequences of the infection are hepatitis, pancreatic necrosis, myositis, myocarditis, uveitis, dermatitis, and encephalitis with the worst lesions being in kittens with congenital infections (Lappin, 2000; McConnell et al., 2007). Analogously with humans, cats with immune deficiency syndrome (e.g., feline immunodeficiency virus) are found to have a predisposition to systemic toxoplasmosis (Davidson et al., 1993; Levy et al., 2004).
In cases of complicated ocular disease or nervous system symptoms, it is always opportune to include a laboratory test for Toxoplasma (Levy et al., 2004). Ocular infections give rise to retinochoroiditis, uveitis with mydriasis, and photophobia leading to blindness. When checking for clinical signs of the nervous system, the veterinarian should take note of any altered motor coordination: signs of hyperesthesia, behavioral changes (e.g., moving in circles with ears lowered, typical signs of fear or aggressiveness), difficulty in mastication or swallowing, epileptic type convulsions, and urinary incontinence (Levy et al., 2004). Various studies have been carried out in the United States on the causes of uveitis in cats and the most common causative agents are T. gondii (>70%), feline leukaemia virus (about 6%), feline immunodeficiency virus (>20%), infective peritoneal virus (about 6%), and mycosis (Lappin, 2000).
The oocysts shed by cat in feces (millions of oocysts after 1–3 weeks from ingestion of sporulated oocysts or tissue cysts) are very resistant in the environment where they sporulate after 1–5 days, thereby becoming infective for humans and the other vertebrates. The oocyst shedding phase is followed by development of immunity in the cat for a variable period of time, lasting for up to >6 years. The relatively short oocyst shedding phase and the length of immunity explain the frequent negative results of the coprologic investigation in seropositive cats (Lappin, 1996; Dubey et al., 2009).
After ingestion of the sporulated oocysts or tissue cysts, T. gondii penetrates the epithelial cells and spreads through the organism via the lymphatic and haematological systems. When there is a cell-mediated and humoral immune response, dissemination of the parasite in tissue does not give rise to evident clinical signs and the tissue cysts develop and persist for years after the infection. Brain, liver, lungs, skeletal muscles, and eyes are the sites of the first phases of multiplication and chronic infection (Greene, 2006). Sometimes the parasite goes through rapid phases of multiplication for no known reason, followed by the appearance of clinical symptoms (Bowman et al., 2002).
Humans
The infection is the direct consequence of eating food contaminated by oocysts eliminated by felids or transported by arthropods, mollusks, or coprophagous birds; by ingestion of cysts or pseudocysts in meat; by inhalation or inoculation of zoites; or by accidental ingestion (hands to mouth for example) of zoites eliminated in secretions and excreta of infected animals (placentas from infected abortions for example) or contained in Flügge droplets (Jones et al., 2009a).
Infection in adults rarely causes disease, except in immunocompromised patients (Mele et al., 2002). Mention should be made here of recent cases of pneumonia attributed to strains present in the past in South America, the so-called genetically atypical strains that are highly virulent even in immunocompetent individuals (De Salvador-Guillouët et al., 2006; Leal et al., 2007).
Prenatal infection is the direct consequence of a primary infection of the mother during pregnancy. Congenital toxoplasmosis ranges from sub-clinical forms to extremely serious cases leading to fetal or neonatal death. In these cases the central nervous system and the eyes are constantly infected, whereas other organs such as the liver, spleen, kidneys, and lungs are rarely involved. Infections in the first stages of pregnancy can bring about abortion, death, or serious fetal damage, such as retinochoroiditis, endocranial calcification, hydrocephaly, and microcephaly (Jones et al., 2009b). In the later stages of pregnancy, on the other hand, T. gondii infections are sub-clinical, even though retinochoroiditis and neurological disorders are sometimes found (Bossi and Bricaire, 2004). Many reports confirm that the classic clinical manifestations (Sabin's tetrad—retinochoroiditis, intracranial calcification, hydrocephaly, and central nervous system abnormalities) are found in 5% of infected newborns, whereas in the majority of prenatal infections there is no congenital disease. Later in life, however, the disease can be re-activated due to persisting tissue cysts (SYROCOT, 2007).
There are no treatments able to eliminate these cysts from an infected organ. This is why some experts suggest that pregnant women with seroconversion documented after conception should be treated with spiramycin (Jones et al., 2009b).
Ocular toxoplasmosis is a consequence of prenatal infection only in one-third of cases and it is considered a probable consequence of postnatal infections (SYROCOT, 2007). The severity of ocular lesions depends on the length of infection and inflammatory intensity. The clinical picture presents necrotizing retinitis with variations in lesion size, number, and aspect. Lesions can be either monolateral or bilateral, with re-activation occurring in 80% of cases. More rarely, but not less serious, are the manifestations of anterior uveitis, and inflammation of sclera and papilla (Hall et al., 2009).
Recent epidemiologic studies indicate that infectious agents may contribute to some cases of schizophrenia. In particular, several epidemiological, behavioral, and neurochemical studies suggested the existence of an association between schizophrenia and past history of infection by T. gondii (da Silva and Langoni, 2009; Dion et al., 2009; Gaskell et al., 2009; Xiao et al., 2009; Yolken et al., 2009; Zhu, 2009).
Food-producing animals
The clinically manifested disease in food-producing animals is not a veterinary health problem, due to its low prevalence (Dubey and Jones, 2008). On the other hand, food-producing animals may represent a real risk for transmission of the disease to humans, either directly or through farming (Samra et al., 2007; Cenci Goga, 2009).
Pigs
Clinical signs of the infection are rare in pigs but can cause premature births and pneumonia. Rare cases of myocarditis and encephalitis have been reported (Dubey and Jones, 2008). Old reports cite nervous system clinical signs (tremors and ataxia), coughing, diarrhea, and a 50% mortality rate, as well as still born and premature births, and neonatal deaths (Farrell et al., 1952; Sanger and Cole, 1955). These reports, however, could have attributed cases to Toxoplasma infection when in reality they were complicated by other factors (Dubey and Beattie, 1988). Probably, the only case of well-documented infection followed by acute toxoplasmosis was that described in 1979 when autopsy on a 4-week-old pig, after 14 days of diarrhoea, revealed signs of lymphadenitis, pneumonia, enchepalitis, and necrotising enteritis, and tachyzoites were detected in all lesions (Dubey et al., 1979). Most infections are actually sub-clinical or feature nonpathognomonic signs such as hyperthermia, anorexia, and tachypnea (Anyarat et al., 2006; Poljak et al., 2008). An attempt at making a summary of available data was made by Tenter et al. in Europe (Tenter et al., 2000) and by Dubey and Jones in the United States (Dubey and Jones, 2008). Seroprevalence ranged from 0% to 64% in fattened pigs and from 3% to 31% in breeding females in Europe, whereas in the United States low prevalence was noted in companies with better management protocols (cat and rodent control). Overall, it can be said that lower prevalence (in many regions almost at zero) is found in indoor stalls with excellent management, whereas higher values, in certain cases above 60%, are found on farms where animals are not kept in enclosed and controlled spaces (for example, on free-range farms or badly managed farms) (Dubey and Jones, 2008).
Cattle
There have been no confirmed cases of clinical toxoplasmosis in cattle and probably many cases of abortion were attributed to T. gondii before the discovery that Neospora caninum can provoke abortions in cattle (Dubey et al., 1988). Bovine are not an important reservoir of the parasite, and even though they can be infected by ingestion of T. gondii oocysts, within a few weeks the bovine host responds and reduces the parasite to unidentifiable levels in laboratory diagnostics (Dubey, 1983, 1986). Studies carried out in the 1990s demonstrated how difficult it is to diagnose the infection in cattle, even with biological testing (Dubey, 1992; Dubey and Thulliez, 1993).
Poultry
Reported clinical cases are very rare (Dubey et al., 2007; Goodwin et al., 1994) with the most recent describing nervous system symptoms in free-range chickens in a family run farm (Dubey et al., 2007). The autopsy on one animal showed necrosis, perivascular lymphocyte cuffs, and gliosis as well as tissue cysts and tachyzoites in the lesions (Dubey et al., 2007). In free-range animals, seropositivity is higher (da Silva et al., 2003; Dubey et al., 2004b; Lehmann et al., 2006).
Sheep and goats
The prevalence of T. gondii in adult sheep and lambs is high and the parasite is known to cause abortions and neonatal mortality in sheep (Samra et al., 2007). Lambs that survive congenital infections grow regularly and are therefore can be a source of infection for humans. A study done at in the late 1990s revealed the parasite in various tissues of lambs slaughtered 7 months after surviving a neonatal infection (Dubey and Kirkbride, 1989). In goats, apart from abortions and neonatal mortality, clinical signs may be present and the parasite can be found in organs and tissue (mainly liver, kidneys, and brain) (Dubey and Beattie, 1988).
Horses
Even though infection is possible in horses (Tassi, 2007), the complete absence of reported evident clinical disease must be mentioned.
Other animals
Sporadic cases of clinical toxoplasmosis have been reported in rabbits, squirrels, canaries, and finches. For squirrels a differential diagnosis from rabies is necessary due to the similarity of symptoms: anorexia, lethargy, taste deprivation, breathing difficulty, and tendency to bite (Salant et al., 2009). In canaries a case of blindness with complete destruction of the eye has been described (Dubey, 2002). The infection has epidemiological importance in deer, bears, and raccoons. Bears and raccoons are omnivores and infection can be explained by the presence of parasites in the intermediate host or the diffusion of oocysts in the environment (Dubey and Jones, 2008). In deer high prevalence can be partly explained by their particular susceptibility especially when living in environments highly contaminated with oocysts (Vikøren et al., 2004; Gauss et al., 2006). Certain animals living in zoos, such as Central and South American monkeys (Platyrrhini, the so-called New World monkeys) (Garcia et al., 2005) and kangaroos are very susceptible to the infection (Miller et al., 1992). Acute toxoplasmosis has been described in lemurs, squirrel monkeys, and marmosets, and has been found to be almost always visceral, followed by cerebral lesions. Lamina propria necrotizing enteritis and lymphadenitis of the mesenteric lymph nodes suggests acquired peroral infection rather than congenital infection (Epiphanio et al., 2003). For some years now, a study is being conducted on the small remaining population of Hawaiian crows (Corvus hawaiiansis) following the death of four birds of toxoplasmosis: a census in 2000 showed there were fewer than 25 specimens remaining (Work et al., 2000).
Population Biology
Recent research on the genetic diversity by highly discriminatory molecular techniques of T. gondii has shown that the parasite is more genetically diverse and not clonal as previously suspected (Lehmann et al., 2006, Dubey and Su, 2009). Intercontinental differences among T. gondii strains have been detected by microsatellite analysis (Lehmann et al., 2006) and, more recently, genotyping data from chickens, pigs, lambs, and deer have demonstrated a high-level genetic diversity (Dubey and Su, 2009). Early genotyping studies on T. gondii led to the description of a clonal population with three main lineages related to virulence in mouse (type I, II, and III) (Darde et al., 1988; Tibayrenc et al., 1991; Sibley and Boothroyd, 1992; Howe and Sibley, 1995).
Recent studies have revealed that South American strains are more genetically diverse and comprise distinct genotypes. These differences have been shaped by infrequent sexual recombination, population sweeps, and biogeography. The majority of human infections that have been studied in North America and Europe are caused by type II strains, which are also common in agricultural animals from these regions. In contrast, several diverse genotypes of T. gondii are associated with severe infections in humans in South America (Beck et al., 2009).
Further epidemiological studies are indicated to determine the role of genetic diversity in T. gondii in transmission between species, pathogenicity, and immunological response (Beck et al., 2009). Recently, several authors found diverse lineages in pigs and chickens and postulated recombination and efficient transmission by oocysts (Dubey and Su, 2009; Velmurugan et al., 2009). Dubey et al. detected high prevalence and high diversity of T. gondii in lambs in the United States: these findings are of great importance for public health, as lamb meat can be an important source of T. gondii infection for humans (Dubey et al., 2008b).
Epidemiology and Transmission Pathways
T. gondii can be transmitted by three routes: congenital infection, ingestion of infected tissue, and contaminated food or water.
Congenital infection
Trans-placental infection has been confirmed in <1% of animals and humans. There are no diagnostic tests available to distinguish between the possible routes of infection; hence, assessment of the role of potential sources of infection is complicated (Dubey, 2008).
Ingestion of infected tissues
Due to the varying organotropism of T. gondii in different host species, the number of tissue cysts produced in a certain organ, in particular in skeletal muscles, varies greatly among different animals (Dubey and Jones, 2008). Therefore, not all animals used for human consumption are of the same public health significance, even if infected with T. gondii. Seropositivity of meat-producing animals does not necessarily reflect the risk that those animals pose to consumers. The meat of cattle and buffaloes rarely contain tissue cysts, although in some areas >90% of these animals are seropositive for T. gondii, whereas seropositive pigs, sheep, and goats can be assumed to harbor large numbers of tissue cysts in their meat (Dubey and Jones, 2008). Tables 1 –5 summarize data for detection of T. gondii cists from animal tissues.
Slaughterhouse from 1 farm.
Pork meat
Only one case of an outbreak after consumption of under-cooked pork has been reported in the literature (Choi et al., 1997). Due to the introduction of intensive farming, the infection has been practically eliminated in many countries. For this reason pork meat is no longer a major source of infection as in the past. However, the present trend to the so-called organic animal raising, or animal-friendly farming, needs to be kept in mind as it could lead to a recrudescence of infection in pigs (Kijlstra et al., 2004). Recent trends in consumer habits, in fact, indicate a shift toward consumption of animal-friendly or organic pigs, which include increased exposure of the pig to the environment, and this will lead to an increased risk of T. gondii in products from such animals (Hill et al., 2010).
Viable T. gondii organisms have been isolated from tissues of pigs collected from abattoirs or farms and the success of isolation varied, in part due to bioassay procedures (Dubey et al., 2002; Dubey, 2009b). In a comprehensive study, the prevalence of T. gondii was determined in 2,094 meat samples each of pork, beef, and chicken, obtained from 698 retail meat stores from 28 geographic areas of the United States. Overall, the prevalence of viable T. gondii in retail pork was very low (0.3%) (Dubey et al., 2005).
Lamb, mutton, and goat meat
Sheep and goats are the main source of meat in Southern European countries (Anonymous, 2007). This is of particular importance because tissue cysts have been found in many edible parts of sheep (Dubey and Kirkbride, 1989), and small ruminants are important in both milk and meat production throughout the world. Consumption of undercooked lamb meat is an important risk factor, as demonstrated by many case–control studies (Anonymous, 2005). Seropositive sheep can be assumed to harbor large numbers of tissue cysts in their meat (Dubey and Jones, 2008). Animal such as sheep and lambs are usually kept in pasture, and therefore they have an increased risk of infection due to contamination of the environment with sporulated oocysts. Sheep and lambs are particularly vulnerable and if the environment is heavily contaminated with oocysts, seroprevalence can exceed 90% (Tenter et al., 2000; Samra et al., 2007). Considering that goats are an important source of food in developing countries, the consumption of goat meat, milk, and cheese products can play a important role in transmission of the disease (Samra et al., 2007).
Beef
Some case–control studies have demonstrated that consumption of under-cooked beef is a risk factor for T. gondii in humans (Baril et al., 1999; Cook et al., 2000). Although seroprevalence in cattle is very high (antibodies are detected in up to 92% of cattle and up to 20% of buffaloes) tissue cysts are found only rarely in beef or buffalo meat (Tenter et al., 2000).
Poultry meat
The parasite has been detected in meat from up to 80% seropositive chickens (da Silva et al., 2003; Lehmann et al., 2006) and T. gondii is expected to be detected more commonly in free range chickens as opposed to intensively housed chickens (Dubey, 2009a; Dubey et al., 2005). Parasite studies in muscle samples (breast) constantly give negative results even in the presence of seropositivity, in animals originating from intensive farming (Dubey et al., 2007).
Horse meat
In a recent study in Italy the parasite was detected in 90% of meat samples (Tassi, 2007). In some countries horse meat is eaten raw and could play an important role in the epidemiology of T. gondii (Tassi, 2007).
Game meats
EFSA data estimate that >50% of the game meat eaten in Europe contains T. gondii cysts (Anonymous, 2007, 2009a). Meat from black bear and deer hunted in the United States had a percentage of positivity ranging from 9.8% to 70% (Dubey et al., 2004a, 2008c; Lindsay et al., 1991).
Milk
Tachyzoites of T. gondii have been detected in the milk of several intermediate hosts, including sheep, goats, and cows (Tenter, 2009). Acute toxoplasmosis in humans has only been associated with consumption of unpasteurized goat's milk (Riemann et al., 1975; Sacks et al., 1982; Skinner et al., 1990; Meerburg et al., 2006). A recent report on how sheep can eliminate T. gondii in their milk is of interest (Fusco et al., 2007). In the past, it has often been thought that the risk of acquiring an infection with T. gondii by drinking cow's milk, if any, is minimal, but it cannot be excluded that any type of milk is a potential source of infection if consumed raw (Tenter et al., 2000).
Eggs
There are discrepant findings in literature regarding the presence of T. gondii in eggs of poultry (Dubey, 2009a). An early study reported that T. gondii tachyzoites may be isolated from raw chicken eggs laid by hens with experimentally induced infection (Jacobs and Melton, 1966), whereas other studies demonstrated very low level or absence of viable organisms in eggs laid by hens experimentally infected (Boch et al., 1966; Biancifiori et al., 1986). Raw hen eggs are therefore unlikely to be a source of infection for humans (Dubey, 2009a).
Water and contaminated food and soil
Several studies have confirmed a link between toxoplasmosis outbreaks and water contamination with oocysts (Kapperud et al., 1996; Bowie et al., 1997; Baril et al., 1999; Cook et al., 2000; Dubey, 2004; Montoya and Liesenfeld, 2004; Kijlstra and Jongert, 2009). The first report dates from 1982, when authors described a waterborne outbreak in U.S. military training in Panama (Benenson et al., 1982). Outside Europe and the United States, water is an important risk factor for infection (Petersen et al., 2009): in 1995 a municipal water system that used unfiltered, chloraminated surface water was the likely source of the largest and best documented community-wide outbreak of toxoplasmosis (Bowie et al., 1997). Sources other than meat and water have been identified: contact with soil (Cook et al., 2000), eating unwashed raw vegetables or fruit (Kapperud et al., 1996), and geophagia in preschool-aged children (Stagno et al., 1980). Infrequent washing of knives after preparation of raw meat was associated with an increased risk of primary infection during pregnancy (Kapperud et al., 1996).
Risk factors for infection in humans and prevention strategies
The most important risk factor for T. gondii infection was found to be under-cooked meat (lamb, beef, and game). However, even true vegans can contract T. gondii infections, confirming the role of oocyst ingestion as a source of infection (Kapperud et al., 1996; Baril et al., 1999; Cook et al., 2000). A multivariate analysis linked the risk of T. gondii infection with eating raw ground beef and rare lamb, eating locally produced cured, dried, or smoked meat, working with meat, drinking unpasteurized goat's milk, and having three or more kittens (Jones et al., 2009a). In one European multicenter study, between 30% and 63% of infections were attributed to consumption of undercooked or cured meat products and 6%–17% to soil contact (Cook et al., 2000).
Available guidelines (Lopez et al., 2000; Leroy et al., 2005) emphasize food hygiene, especially thorough cooking of meat and meat products, and reduced contact with soil and cat feces. Given that freezing meat for at least 2 days at temperature below −12°C can kill tissue cysts (Kotula et al., 1991), people at risk should be encouraged to buy frozen meat. Tissue cysts are also killed by heat treatment: internal temperature of 56°C for 10 minutes in sufficient to guarantee a Toxoplasma-free meat (Dubey et al., 1990). Tachyzoites are killed by pasteurization and heating; therefore, it is advisable that milk, in particular goat's milk, should be pasteurized or boiled before human consumption (Tenter et al., 2000). Any type of cooking would kill tachyzoites in eggs (Tenter et al., 2000).
Several epidemiological studies have identified the risk of T. gondii infection at farm level and preharvest prevention has been postulated (Kijlstra and Jongert, 2008, 2009). This includes strict rodent control (especially in the pig farming, where consumption of rodent remnants could be a direct source of infection for pigs) and certification that the farm and surroundings are free of oocyst-shedding cats (Kijlstra and Jongert, 2009). The use of sterilized feed and bedding along with control measures to prevent the access of pests has led to a Toxoplasma-free production system in certain pig and broiler farms (Dubey and Jones, 2008).
Practicing small animals veterinarians are frequently asked to diagnose toxoplasmosis in cats because of the potential public health risk and clinical illness. The clinical diagnosis of Toxoplasmosis is dependent on a thorough understanding of the life cycle of the parasite and its relationship to mammalian hosts. Pregnant women, AIDS victims, and other immunosuppressed patients commonly ask their veterinarian about when a cat has previously shed oocysts and when will shed oocysts again in future. While fecal examination is an adequate procedure for detecting current oocysts shedding, it fails to predict whether a cat has shed oocysts in the past or will shed in the future (Schares et al., 2008). On the other hand, there is no serological assay that can accurately predict when oocysts were shed in the past (Dabritz et al., 2007). For these reasons veterinarians should be prepared, in terms of both scientific knowledge and communication skills, to educate cats owners and to encourage responsible cats ownership. Key points, yet not easily accepted by all cats lovers, are spaying and neutering cats to control the size of owned cat population, keeping cats indoor, and collecting feces in litter boxes for disposal. Cat feces should not be disposed in toilets because oocysts survive treatments for raw sewage (Wainwright et al., 2007a, 2007b).
From a public health perspective, the best approach would be to vaccinate cats. From the beginning of the 1990s some experiments based on administration of attenuated live bradyzoites to cats have been completed with success (Frenkel et al., 1991; Mateus-Pinilla et al., 1999). At the moment research in this sector is languishing due to lack of interest on the part of health authorities and therefore a lack of financing. Difficulty in producing the vaccine (a patent exists, U.S. Patent 5045313, filed on July 7, 1989), based on the multiplication of bradyzoites in mice followed by purification, and a limited economic incentive for the pharmaceutical industry could be overcome by the use of cell cultures or recombinant vaccines (McAllister, 2005).
Laboratory Diagnostics
Serological testing
Fluids that can be sampled include those present in the thoracic cavity of aborted fetuses, milk, and extracts of infected tissue or liquid drained from muscles that have been frozen and then thawed. The plethora of commercially available methods used in various laboratories makes it difficult to compare results, which is actually the most important aspect today to control the spread of the disease (Fricker-Hidalgo et al., 2009).
Modified agglutination test, Immunofluorescence antibody test, enzyme-linked immunosorbent assay, and Sabin-Feldman dye test
Modified agglutination test (MAT) is the most commonly used test today for pets and wild animals due to its ease of use and safety (Shaapan et al., 2008). Agglutination uses tachyzoites inactivated with formalin and does not require species-specific reagents (Shaapan et al., 2008; Macrì et al., 2009). A study revealed that MAT has the highest sensitivity, followed by enzyme-linked immunosorbent assay (ELISA), whereas among serological tests, immunofluorescence antibody test (IFAT) showed the lowest sensitivity. On the other hand, IFAT had the highest specificity, followed by MAT and ELISA (Shaapan et al., 2008).
IFAT is the simplest test and can be easily performed even by inexpert lab technicians. IFAT also uses tachyzoite antigens treated with formalin, but species-specific conjugates are necessary (Shaapan et al., 2008). The variability of the sensitivity and specificity according to the T. gondii strain employed, and subjectivity in interpreting the fluorescence reaction make it almost impossible to compare results from different laboratories. This is the reason for the numerous data in the literature that show extreme variability even in the same regions and in the same type of farm (Shaapan et al., 2008; Macrì et al., 2009). A recent study has revealed that there is an extremely low proportion of accordance between IFAT and MAT in cat and dog sera (Macrì et al., 2009).
Many kits, based on ELISA, are commercially available to find serum antibodies in different animal species. The methodology can be automated and this makes it attractive for use in large-scale epidemiological studies. For the most recent tests introduced on the market, manufacturers declare sensitivity and specificity greater than IFAT and other serological tests (Hosseininejad et al., 2009). ELISA tests to demonstrate IgG to diagnose active infections are available in the market although there is not full agreement among researchers on their sensitivity and specificity (Shaapan et al., 2008; Hosseininejad et al., 2009). Recently, an indirect ELISA test with purified tachyzoite surface antigen SAG1 has been proposed (Hosseininejad et al., 2009). SAG1 antigen is stage-specific and only detectable in the tachyzoite stage (Pietkiewicz et al., 2007).
Sabin-Feldman dye test is based on the inability of tachyzoites to absorb the dye when, tested with anti-Toxoplasma anti-serum, the immunocomplex has been formed. The test is highly sensitive and specific, has been used on humans, and can be adapted to different animal species, except for bovines where a globulin can give false-positives. The method requires live tachyzoites, is expensive, and is restricted to use in specialized laboratories and expert lab technicians (Ozkan et al., 2008).
IgG versus IgM
Serological tests, for example, ELISA, can diagnose active infection (positive IgM and negative IgG tests) (Hosseininejad et al., 2009). The Sabin-Feldman dye test, in cases of seroconversion (or increase in titer fourfold in 2–3 weeks), allows diagnosis of active infections (Bowman et al., 2002). MAT is an ideal test in veterinary healthcare settings to detect IgG because it does not require species-specific conjugates used in serologic assays based on the immunofluorescent antibody and ELISA methods. On the other hand, performance of the MAT for detection of IgM antibodies against T. gondii in naturally infected animals is less clear and there have been few systematic evaluations of the assay for this purpose. IgM antibodies can be detected 1 to 2 weeks before the development of a measurable IgG antibody titer (Svobodova et al., 1998). The primary problem with testing for anti-Toxoplasma IgM is lack of specificity (Lappin, 2010). In general, for animals with a positive IgM result and negative IgG on repeated follow-up examinations, T. gondii can be ruled out as potential etiology, whereas the detection of T. gondii–specific IgG antibodies on the follow-up examination conducted within 3 weeks of the initial IgM titer retrospectively confirms that the animal is infected with T. gondii parasites (Lappin, 2010).
Other tests
Tachyzoites and tissue cysts can be found in tissue sections and organs fixed and stained with hematoxylin and eosin and immunohystochemistry. Since tissue cysts or tachyzoites are not evenly distributed in the tissues and the body (van Maanen et al., 2004), there is a low correlation between immunohistochemistry and serological tests (Moré et al., 2008).
Another diagnostic tool is the cultivation of T. gondii in cell culture. However, the need for adequate laboratories and technical experience makes it a little-used routine diagnostic tool. The sensitivity of this test is also one of the lowest especially for samples abused during storage and transportation (James et al., 1996).
Polymerase chain reaction is used for the detection of the DNA of the parasite in blood, fluid, and tissues: the specificity of this test is almost 100%, but the difficulty to extract DNA and concentrate large sample quantities results in limited sensitivity (Alfonso et al., 2009). It should be considered also that cyst distribution is random and that parasite density in the cysts could be very low (Piergili Fioretti, 2004; Alfonso et al., 2009).
The present trend, for obvious ethical reasons and for animal wellness, is to limit or avoid the use of biological testing for diagnosis. However, the possibility of inoculating or feeding Toxoplasma-free laboratory animals (mice and cats) to later demonstrate T. gondii in organs and tissues should be held in consideration, especially for the high sensitivity and specificity that, unfortunately, make this biological testing the gold standard (James et al., 1996). Other tests include immunochromatographic assay, carbon immunoassay, direct agglutination test (which is also sold in kits and is used in wild animals as it does not require species-specific conjugate), latex agglutination test, and indirect hemagglutination test. Immunoblotting, even though it cannot be considered a routine test, is extremely important to study serum reactivity to many T. gondii antigens (Anonymous, 2007).
Footnotes
Acknowledgments
The authors thanks all those colleagues who have helped in our ongoing research projects, in particular Prof. Barend Louis Penzhorn and Dr. Nada Abu Samra of the University of Pretoria, School of Veterinary Medicine, Onderstepoort, and Dr. Carlo Crotti and Dr. Ludovico Renda veterinary practitioners in Perugia, Italy. The authors also thank Dr. Marcello Tosti for providing the photograph and express sincere appreciation to members of Polyglot, Perugia, for a careful reading and comments on the article. The findings and conclusions in this review are those of the authors and do not necessarily represent the views of the University of Perugia, the University of California, or the University of Pretoria.
Disclosure Statement
No competing financial interests exist.