
Abstract
Mexican fruit fly Anastrepha ludens Loew (Diptera: Tephritidae) is one of the most important insects infesting fruits. Although high pressure has been proposed as an alternative quarantine process for this pest, conditions applied to destroy eggs and larvae can also damage the fruits. The objective of this study was to assess the biological viability of A. ludens eggs treated by high-pressure processing at 0°C, establishing whether nondestroyed eggs and larvae preserved their ability to develop and reproduce. One-, 2-, 3-, and 4-day-old eggs were pressurized at 50, 70, or 90 MPa for 0, 3, 6, or 9 min. The hatching ability of pressurized eggs and their capacity to develop larvae, pupae, and reproductive adults were evaluated. The ability of pressurized larvae to pupate was also registered. The results showed that most of the eggs that resisted the treatments were able to produce adults with capability to reproduce a new generation of A. ludens flies. Larvae were less resistant than eggs. All larvae were killed at 90 MPa for 9 min. The pressure induced the expression of heat shock proteins (HSP) in second- and third-instar larvae. The HSP showed a baroprotective effect in A. ludens larvae. These results are relevant to the industry because they show that eggs of insects infesting fruits treated by high-pressure processing were able to pupate after treatments. This indicates that efforts must be addressed to destroy all eggs because most of the surviving organisms could reach an adult stage and reproduce.
Introduction
Cold storage has been used as a quarantine method since the beginning of the 20th century (Mangan and Hallman, 1998). USDA-APHIS has approved holding fruit at 1.1°C, or less, for 12 days as a cold treatment to be used against this species for 17 commodities from 49 countries (Phillips et al., 1997; Gazit et al., 2004).
Although cold treatment was approved as a quarantine method, it has been reported that it can reduce the commercial value and quality of many tropical fruits because of chilling injury (Phillips et al., 1997; Gazit et al., 2004; Lanza et al., 2005), which imposes a practical limitation to select cold treatment as a quarantine method (Lederman et al., 1997; Mangan and Hallman, 1998).
The use of high-pressure processing (HPP) has been reported as an alternative quarantine process for Mediterranean fruit fly, Mexican fruit fly, western cherry fruit fly, and codling moth (Butz and Tauscher, 1995; Neven et al., 2007; Candelario-Rodríguez et al., 2009; Velazquez et al., 2010a, 2010b); however, early studies have found that the HPP conditions required to destroy the eggs and larvae of these pests caused unacceptable damage to the fruits.
The use of combined treatments to meet quarantine requirements has been reviewed for a number of combined approaches (Mangan and Sharp, 1994). Using cold temperatures combined with HPP has been reported as an effective alternative to destroy eggs and larvae of Mexican fruit fly (Hernandez-Ortiz, 1992; Velazquez et al., 2010b). However, pressurization levels above 100 MPa can induce physical and physiological changes in the targeted fruit (Neven et al., 2007; Candelario-Rodríguez et al., 2009). On the other hand, it is important to consider that profitability of HPP requires short processing times (Velazquez et al., 2005a, 2005b). Therefore, the operating conditions of HPP to destroy eggs and larvae should be less than 100 MPa and short times (less than 10 min).
On the other hand, heat shock proteins (HSP) act in the organisms as molecular chaperones (Craig et al., 1993). These proteins protect the organisms from injury at high temperatures by avoiding protein denaturation. HSP appear to be capable of recognizing simple structural motifs shared by many unfolded proteins (Denlinger and Yocum, 1998). HSP have been reported to be expressed during high-pressure treatments of yeast and bacteria, protecting microorganisms against pressure damage. However, it has not been reported in fruit fly as a response against pressure.
The objectives of this work were to determine the combined effect of HPP and cold temperature (0°C) on the capability of Mexican fruit fly eggs and larvae to survive and preserve their biological capacity to develop and reproduce and, second, to elucidate whether HPP can induce the production of HSP in Mexican fruit fly as a possible protecting mechanism.
Materials and Methods
Sampling site
The experimental work was conducted at the facility of Moscafrut in Metapa de Domínguez, Chiapas, México, which produces Anastrepha ludens for sterile insect release programs. The eggs and larvae used in this study were viable (nonsterilized), supplied by this facility, and reared as reported elsewhere (Dominguez et al., 2010).
High-pressure system
Pressure treatments were carried out as reported elsewhere (Candelario et al., 2010). Briefly, a mixture of 5:1 water with crushed ice:anticorrosive lubricant (Hydrolubric 120-B; EF Houghton, Valley Forge, PA) was used as pressurizing fluid to maintain a temperature near 0°C during treatments. The temperature of the fluid was never higher than 0°C as the ice was still present at the end of the treatments.
Eggs and larvae were pressurized at 50–90 MPa for 0–9 min at 0°C. The time required to reach process pressure was less than 44 sec and the release time was always less than 30 sec. A treatment for 0 min is a treatment wherein the pressure is reached and released immediately.
Larvae for studies on polyacrylamide gel electrophoresis (see below) were treated at 50–150 MPa for 0–20 min.
High pressure treatments on eggs
One-, 2-, 3-, or 4-day-old eggs were processed independently in two runs. A 4.5 mL aliquot of an egg/water solution containing 24,200 eggs/mL (108,900 total eggs) was taken from a 10-L plastic container and placed into three Eppendorf® tubes with 2 mL capacity. The tubes were filled completely with water, closed immediately, and pressurized.
High-pressure treatments on larvae
The same high-pressure treatments were applied to 30,000 larvae of each instar. Briefly, larvae were manually separated, weighed, and counted using a small quantity of talc powder to facilitate handling. The number of larvae in 5 g (by triplicate) was used to estimate the amount required to obtain 30,000 larvae. Larvae were introduced into plastic pouches of 200 mL and sealed immediately before the high-pressure treatment. After treatment, the surviving larvae were transferred in groups of 100 larvae to Petri dishes containing vermiculite and held at 20°C and 80% relative humidity in the dark for 48 h. After the holding period, the number of pupae formed was recorded. The survivorship to emergence or reproductive capacity of the flies was not registered.
Determination of eggs hatching
After pressurizing treatments, the eggs from one Eppendorf tube were extracted using a brush and arranged in three rows of 100 eggs in Petri dishes using a Zeiss Stemi SV 6 stereoscopic microscope and incubated at 26°C ± 1°C for 7 days to monitor egg hatching. Three replicates of this bioassay were performed.
Determination of surviving eggs' capability to form larvae, to pupate, and to produce adults and the reproductive capacity of emerged adults
These determinations were performed as reported elsewhere (Candelario et al., 2010).
Microphotography of eggs
Nonhatching eggs were photographed using a digital camera (Nikon DXM1200F) attached to a dissecting microscope (Nikon SM21500).
Electrophoresis
One-dimensional sodium dodecyl sulfate–polyacrylamide gel electrophoresis was performed according to Laemmli (1970). In all cases, a 4% stacking gel and a 12% separating gel were used. Electrophoresis was performed at 20 mA per gel. An aliquot of molecular weight (MW) standard mixture (MW.SDS.70L; Sigma Chemical) was included in all gels.
Statistical analysis
Statistical analysis was performed using a Statgraphics 5.0 (Software Publishing Corporation, Bitstream, Inc.). In egg's bioassay, LSD's multiple range tests were used to determine significant differences (p < 0.05) between treatments.
Results
Effect of high pressure on eggs
The effect of the pressurizing level and treatment time is shown in Table 1. The hatching ability of eggs was affected by both variables, time and pressure level.
Mean values and standard deviations of two studies with two replicates are shown.
Different letters in the same row indicate significant differences (p < 0.05) between time of processing for each level of pressurizing.
The hatching ability of 1- and 2-day-old eggs was more sensitive to this treatment than the 3- and 4-day-old eggs. All 1-day-old eggs completely lost their ability of hatching by using 70 or 90 MPa for 3 and 6 min. However, some of the 3-day-old eggs were able to hatch even after 90 MPa for 9 min.
Figure 1 shows the effect of HPP on the inner structure of A. ludens eggs that were not able to hatch.

Anastrepha ludens eggs pressurized at 0 (control), 50, 70, or 90 MPa for 9 min at 0°C.
Development of pressurized eggs to form larvae, pupae, and adults
The results for the ability of pressurized eggs to form larvae are shown in Table 2. The rate of larvae formed from pressurized eggs that survived until third-instar stage was affected by time and pressure level. Nonpressurized control eggs showed more than 91% of larvae survival.
Mean values and standard deviations of two studies with two replicates are shown.
The number of eggs that were able to hatch at 50 and 70 MPa for 0 min for 2-day-old eggs (Table 1) were slightly lower than the number of surviving larvae reported in Table 2. This difference was most likely caused because data entries in Tables 1 and 2 represent an approximation obtained by population estimations. Table 1 reports an estimate obtained from the rate of hatching of 300 eggs (by triplicate), whereas data of Table 2 were obtained by counting the number of larvae in 5 g, and such data were employed to calculate the total number of larvae.
The effect of HPP on the ability to pupate of surviving larvae was determined and results are shown in Table 3. Most of the surviving eggs were able to form third instars (Table 2) with enough vitality to develop until the pupae stage was reached (Table 3). The ability to form pupae decreased at a minimum when eggs were pressurized at 90 MPa. Even at 90 MPa for 9 min, some pressurized eggs were able to reach the pupae stage.
Mean values and standard deviations of two studies with two replicates are shown.
Most of the eggs that were able to survive the treatments and preserved their ability to form pupae (Table 3) preserved also their ability to form adults (Table 4).
Mean values and standard deviations of two studies with two replicates are shown.
The adults obtained from pressurized eggs were matched with individuals of their opposite sex to determine whether they retained their reproductive ability and whether the new eggs were able to hatch (Table 5). Flies obtained from pressurized eggs were able to reproduce and their eggs were able to hatch.
Mean values of one study with three replicates are shown.
Effect of high pressure on larvae
Larvae showed very low resistance to HPP even at 50 MPa (Table 6). The surviving rate decreased drastically with increasing time and pressurizing level. Third instars showed the highest resistance to HPP. However, larvae from all instars did not survive when pressurized at 90 MPa for 9 min. A high rate of surviving larvae preserved their ability to pupate (Table 7).
Mean values of one analysis for approximately 30,000 larvae are shown.
Mean values of one analysis for approximately 30,000 larvae are shown.
Expression of HSP
The expression of HSP by pressurized second and third instars is depicted in Figures 2 and 3, respectively. Larvae were pressurized at 50–150 MPa for 0–20 min. Second instars showed the presence of proteins with MW greater than 66 kDa, corresponding to the HSP70 family. HSP of low MW were also expressed in a very low concentration (Fig. 2). Third instars showed a high concentration of the HSP70 family. HSP of low MW were also observed but in lower concentrations.
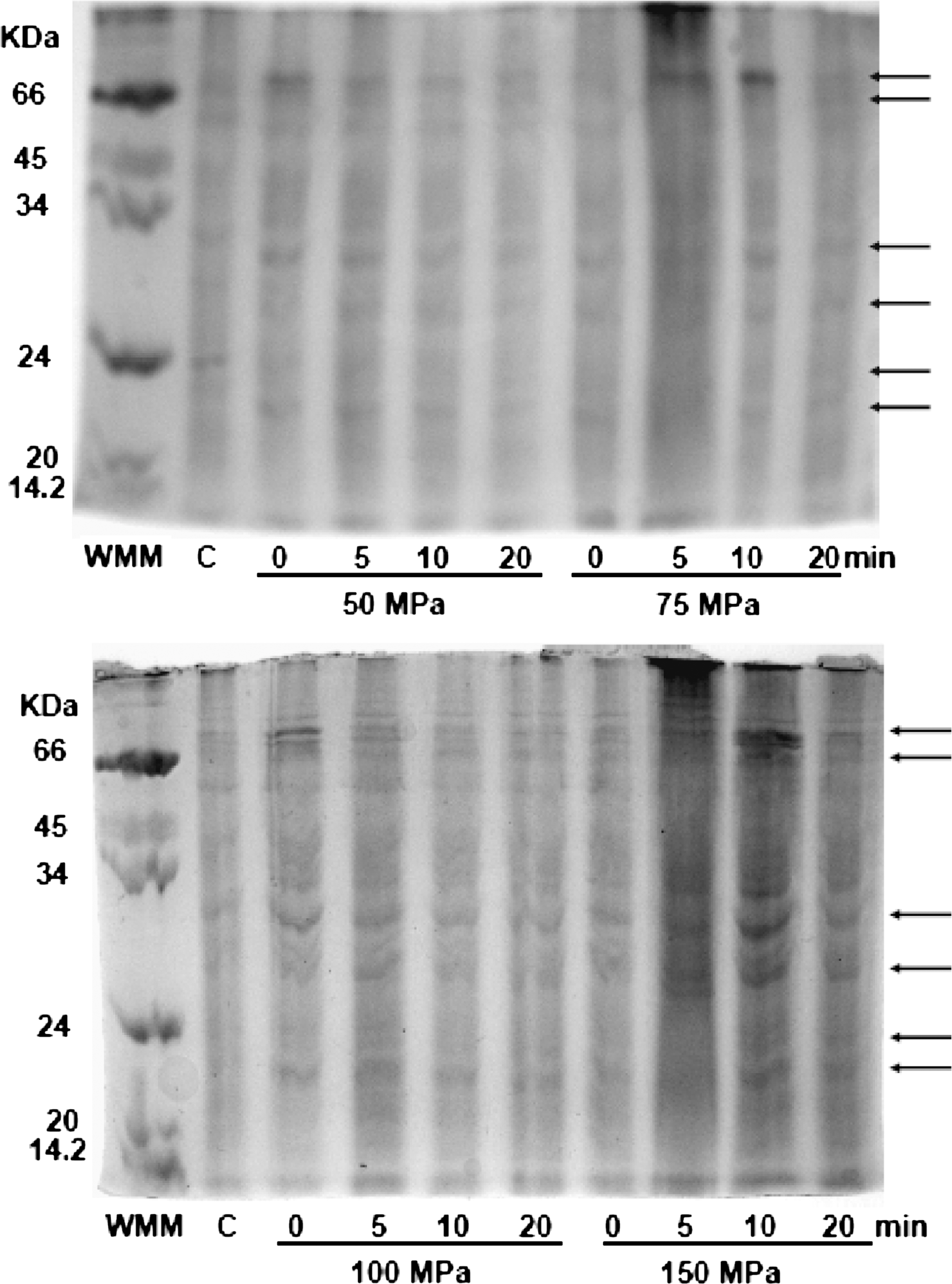
Electrophoretic pattern of proteins extracted from second instars pressurized at different conditions. Arrows indicate the presence of proteins expressed as a result of high-pressure processing. Lane 1 shows the molecular weight marker proteins and lane 2 is a nonpressurized sample (control).
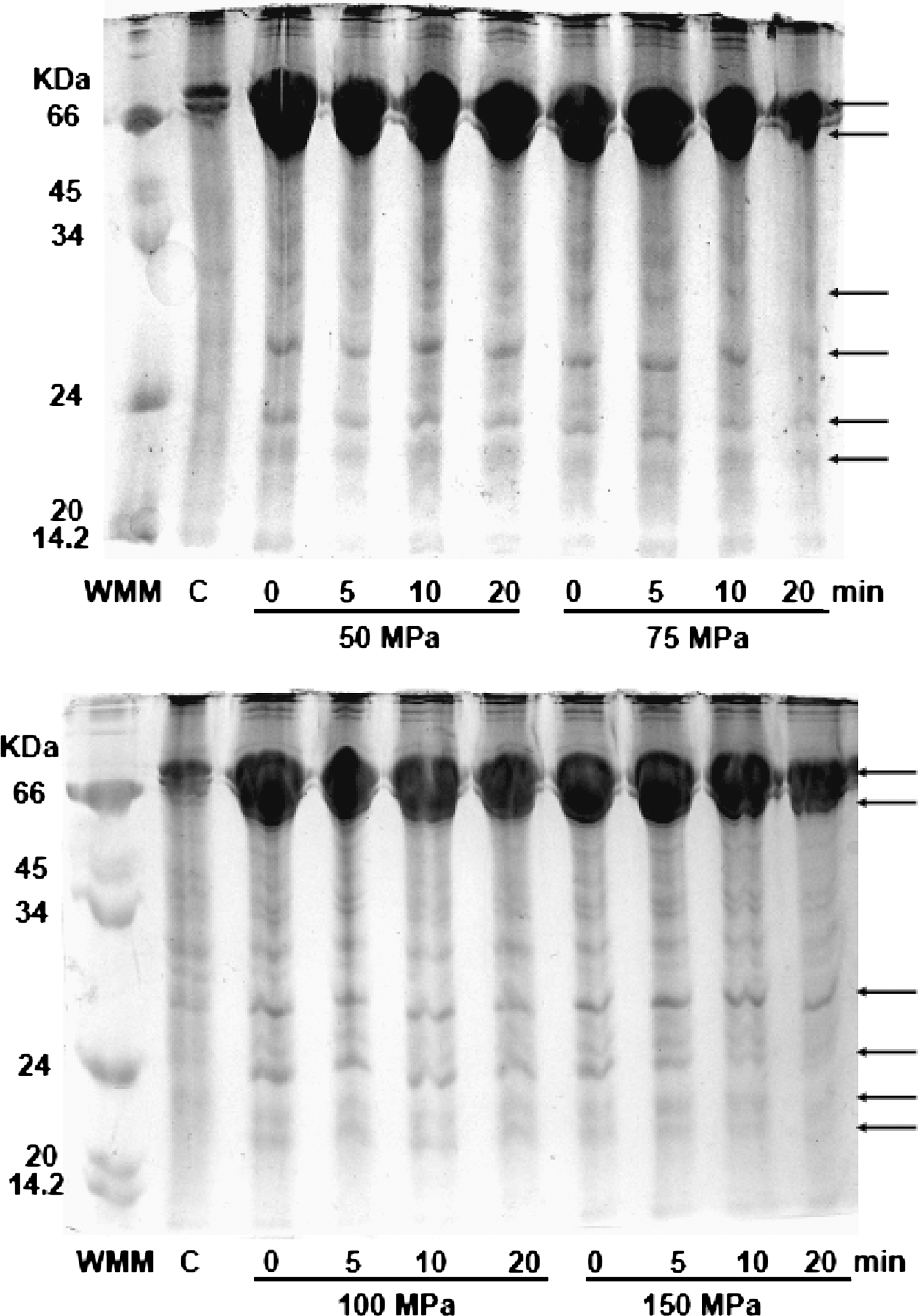
Electrophoretic pattern of proteins extracted from third instars pressurized at different conditions. Arrows indicate the presence of proteins expressed as a result of high-pressure processing. Lane 1 shows the molecular weight marker proteins and lane 2 is a nonpressurized sample (control).
Discussion
Effect on eggs hatching
It was reported that HPP at 50 MPa and 25°C showed a minimum effect on the eggs hatching of A. ludens even after 20 min. The effect of pressurizing at 50 MPa was improved by decreasing the initial temperature to 0°C (Velazquez et al., 2010b). However, in such study, the final process temperature fluctuated between 0°C and 1°C and the hatching level ranged between 22% and 82%, depending on the age of the eggs. In our study, a greater inhibition of eggs hatching was observed at 50 MPa (Table 1), because the temperature of the pressurization chamber was held constant at 0°C during the entire process. These results suggest the importance of temperature control on the lethal effect of HPP.
Some eggs showed higher resistance to HPP and preserved their ability to hatch. This resistance could be associated with the physiology of the eggs, because HPP is instantaneous and uniform (Téllez-Luis et al., 2001; Torres and Velazquez, 2005; Velazquez et al., 2005a, 2005b).
One- and 2-day-old eggs pressurized at 50–90 MPa for 9 min showed a change in the inner structure, compared with the control eggs (Fig. 1). The changes were more evident at higher pressures. Eggs are composed mainly of water and proteins. The effect of HPP on egg hatching has been associated with a change in the structure and permeability of external and internal membranes, denaturation/aggregation of proteins, or a combination of both factors (Borderías et al., 1997; Fernández-Martín et al., 1998; Uresti et al., 2004; Velazquez et al., 2010b).
In this study, it was observed that HPP induced, in 1-day-old eggs, the flocculation and separation of proteinaceous structure, compared with control eggs (Fig. 1). This behavior was more evident when the pressure was increased until a full disruption of the cellular membrane was achieved at 90 MPa for 9 min. A similar behavior was observed for 2-day-old eggs, although these eggs did not show the rupture of the cellular membrane.
The loss of hatching ability in eggs induced by HPP at 0°C could be due to protein denaturation/aggregation, because this phenomenon affects the viability of the organism (Farr, 1990; Pothakamury et al., 1995; Téllez-Luis et al., 2001).
Although most eggs lost their hatching ability, some eggs were able to survive and hatch. This resistance has been reported in some specimens for other quarantine treatments such as cold or heat shock injury as well as long-term heating and chilling injury (Denlinger and Lee, 1998; Denlinger and Yocum, 1998; Ashie et al., 1999; Uresti et al., 2005).
Surviving larvae obtained from pressurized eggs
The rate of surviving larvae reaching the third instar (Table 2) was lower than the percentage of hatched eggs (Table 1). This difference may be associated with physicochemical changes caused by the pressurizing treatment on proteins involved in DNA replication and transcription (Farr, 1990; Pothakamury et al., 1995; Téllez-Luis et al., 2001). Such changes could induce the formation of larvae with physiological deviations, which inhibit their growth and ability to reach third instar.
Formation of pupae from surviving larvae and adults from pupae
Although some larvae were not able to pupate, most of them did (Table 3), indicating that most of the surviving eggs preserved their physiological properties and biological functionality associated with growth. Most of the pupae emerged as adult flies, confirming that most of the surviving eggs preserved their viability.
Viability of eggs produced by adults obtained from pressurized eggs
Most of the adults that emerged from eggs resisting HPP preserved their ability to reproduce and lay viable eggs (Table 5). Recently, it was reported that eggs resisting HPP treatments at initial 50°C maintained their ability to reproduce and lay viable eggs (Candelario et al., 2010). These results confirm that efforts to optimize HPP as a quarantine method for A. ludens must be focused to destroy both eggs and larvae, because the surviving organism could be able to reproduce.
Effect of pressurization on larvae
HPP killed most of the larvae by inducing changes in their external and inner structure as well as by modifying their proteins. The physical damage of larvae by HPP included the presence of dark regions, air bubbles, separation of external membrane, and liquid accumulation (Candelario et al., 2010).
HPP induced the expression of hsp in second and third instars (Figs. 2 and 3). Second instars showed a lower expression of HSP than third instars associated with a lower resistance to HPP (Table 6). The higher resistance of the third instars to HPP seems to be associated with the expression of proteins from the HSP70 family. To our knowledge, there are no reports of the presence of HSP in fruit flies derived from a pressurizing treatment. All organisms respond to heat and a variety of other stresses such as cells anoxia, ethanol, and certain heavy metal ions, by inducing the synthesis of HSP. The stress proteins encoded by the HSP70 and HSP90 gene families in response to elevated temperatures are among the most highly conserved proteins. The HSP themselves or their close relatives are present in all organisms at normal temperatures and play vital roles in normal cell function (Lindquist and Craig, 1988).
HSP70 appears to be the most prominent contributor to thermotolerance in insects. These are the proteins that responds most dramatically to heat shock, and it can be induced more than 1000-fold in response to heat shock (Velazquez et al., 1983; Denlinger and Yocum, 1998). In our study, HSP were expressed, as a response to high pressure, in a high concentration in third instars, thereby protecting them against pressurization and increasing their surviving rate during HPP.
Effect of pressurization on expression of HSP
The small HSP that were less expressed than the HSP70 family, had a modest 10-fold increase in expression but they are important in the high temperature response (Denlinger and Yocum, 1998). During HPP the small HSP were expressed in low concentrations in both second and third instars (Figs. 2 and 3), but seemed to be associated with the resistance of some individuals to HPP. Small HSP with an MW of 15–30 kDa are ubiquitous and conserved. These proteins act as molecular chaperones in folding reactions and play a role in the formation or maintenance of the native conformation of cytosolic proteins. HSP promote the functional refolding of proteins after denaturation.
In Saccharomyces cerevisiae, HSP provided protection against subsequent pressure damage. The parallel induction of barotolerance and thermotolerance by heat shock suggested that high-pressure and high-temperature effects on yeast may be physiologically tightly linked (Iwahashi et al., 1991; Jakob et al., 1993; Aertsen et al., 2004). The data found in our study are in agreement with these results. The HSP showed a baroprotective effect on A. ludens larvae.
Conclusions
Hatching of A. ludens eggs was inhibited by pressurizing at 50–90 MPa for 0–9 min at 0°C. However, some eggs were able to survive the process and preserved the ability to hatch, form larvae, pupate, and develop into adults with reproductive capacity. The larvae of A. ludens were less tolerant to pressure than eggs. All larvae died at 90 MPa for 9 min. These results indicate that efforts must be addressed to destroy all eggs and larvae because most of the surviving organism could be able to reach adult stage and reproduce.
Footnotes
Acknowledgments
The authors are grateful to PROMEP for financial support to this work, to José Manuel Gutiérrez Ruelas, Director de Moscas de la Fruta, DGSV-SENASICA SAGARPA, for logistic support, and to Refugio Hernández and Emilio Hernández (Programa Moscafrut SAGARPA-IICA) for technical and scientific advice.
Disclosure Statement
No competing financial interests exist.