
Abstract
In this study, 20 samples from three different sources of animal feed were investigated and six bacterial isolates were identified. The susceptibility of four natural antimicrobials, namely, eugenol, cinnamaldehyde, thymol, and carvacrol, against six of these isolates was determined. Carvacrol and eugenol showed better inhibitory effects with larger zones of inhibition. The minimal inhibitory concentration for a range of antibiotics on the susceptibility of two isolates (namely, Sphingomonas paucimobilis and Klebsiella oxytoca) was investigated using the VITEK® 2 microbiological identification system. Both isolates showed a variety of resistance to 18 antibiotics. The minimal inhibitory concentration and fractional inhibitory concentration index of those two isolates for ampicillin and nitrofurantoin in combination with four phenolic compounds was determined. Synergistic interactions were found for most antimicrobial/antibiotic combinations; thymol and carvacrol were very effective (fractional inhibitory concentration ≤0.5) in combination with all antibiotics tested against S. paucimobilis and K. oxytoca, respectively. Ultra performance liquid chromatography techniques were used to investigate the physiological effects of the four natural antimicrobials against those two isolates. Two identical peaks were found to be systematically different between cinnamaldehyde-treated and -untreated cells. The identity of the peaks is unknown and further investigation is needed.
Introduction
A
Due to the accumulation of antibiotic-resistant bacteria and genes, leading to the treatment failure or longer convalescence in both humans and animals, research on antibiotic replacement is currently emerging (Ultee et al., 2002; Bagamboula et al., 2004; Pei et al., 2009). Use of natural antimicrobials such as extracts from spices and/or herbs for the purpose of food preservation and/or the substitution of antibiotics has arisen. It has been well documented that essential oils or their active components from wild plants' natural defense systems can act as effective antimicrobials (Ultee et al., 1999; Olasupo et al., 2003; Li et al., 2010). Thus, the aim of the present work was to examine the effectiveness of natural antimicrobials against bacteria isolated from animal feeds.
In this study, we examined the growth inhibitory effect of several natural antimicrobials, namely, eugenol, carvacrol, thymol, and cinnamaldehyde, in relationship to selective antibiotic-resistant bacteria, including Klebsiella oxytoca and Sphingomonas paucimobilis, isolated from animal feeds. An investigation of the minimum inhibitory concentration (MIC) and synergistic interaction between natural antimicrobials and antibiotics to which the bacteria were resistant was then followed. A newly developed ultra performance liquid chromatography (UPLC) method was also used to explore bacterial physiological changes before and after natural antimicrobial treatments to selected isolates.
Materials and Methods
Materials
Three types of animal feed sampled in this study were feed for cattle, ducks, and hogs, which were inspection samples provided by suppliers in the greater Shanghai region. Listeria monocytogenes ATCC 19115 was purchased from ATCC and Escherichia coli isolates were a generous gift provided with the VITEK® 2 system (bioMérieux). Selected natural antimicrobials (substituted phenolic compounds) were known to be active against the bacteria used in this study (Pei et al., 2009). The specific antimicrobial agents tested were eugenol (99% purity), carvacrol (99%), thymol (99.5%), and cinnamaldehyde (>93%). Antibiotics used in this study were ampicillin sodium and nitrofurantoin (Stock and Wiedemann, 2001). The antibiotics and natural antimicrobials as well as p-iodonitrotetrazolium chloride for the Gram-staining assay were purchased from Sigma-Aldrich.
Bacteria isolation and identification
Twenty-five grams of the sample was mixed with 225 mL 0.85% sodium chloride solution (Sigma) in a 250 mL sterilized flask and vigorously shaken. Aliquots of 0.2 mL were serially diluted and enumerated on Tryptone Soya Agar (TSA; Oxiod) in duplicate. The cultures were incubated at 37°C for 24–48 h and selected colonies were subcultured 3–4 times followed by Gram staining (GB/T4789.28-2003; CLSI, 2002). The isolates were identified by the VITEK® 2 microbiological identification system (bioMérieux) and cryogenically stored at −70°C in 40% glycerol solution (Zhang et al., 2010).
Natural antimicrobial susceptibility test
Twenty bacterial isolates from animal feeds were grown in Tryptone Soya Broth (TSB; Oxoid) separately to achieve a ca. 108–109 CFU/mL population and adjusted to 105–106 CFU/mL with a 0.85% sodium chloride solution. Cultures were then spread evenly on TSA with sterile bent glass rods. Four sterilized paper discs (0.6 cm diameter) containing natural antimicrobials ranging from 0.625 to 2.5 mM/mL along with one ethanol control were placed on each plate. Each natural antimicrobial-containing paper disc was prepared by adding 500 μL of natural antimicrobial solution diluted with ethanol (25% v/v) (Sigma-Aldrich) to various concentrations. Fifty paper discs were immersed in solution for 5 h and dried at 37°C overnight before use. Plates were incubated at 37°C for 18–24 h before recording zones of inhibition.
MIC determination
The antibiotic resistance of isolated bacteria was investigated by the VITEK® 2 microbiological identification system by using Gram-negative VITEK susceptibility test cards AST-GN. The MIC values of antibiotics alone and in combination with a pretreatment of 0.1 mM/mL natural antimicrobials were determined. While MIC values of natural antimicrobials alone and in combination were determined by standard broth microdilution (Li et al., 2010) performed in 96-well microplates (Bio-Rad). Natural antimicrobial (0.1 mL)-treated cell suspensions were diluted two-fold in TSB broth in eight separate wells with concentrations of 10, 5, 2.5, 1.25, 0.625, 0.312, 0.156, and 0.078 mM/mL; uninoculated wells served as negative controls. The microplates were sealed with sealing film UC-500 (Sigma-Aldrich) and incubated at 37°C for 18–24 h. Forty microliters of p-iodonitrotetrazolium violet (Sigma-Aldrich) dissolved in distilled water was added to each well with a final concentration of 0.2 mg/mL. The microplates were then incubated at 37°C for 1–2 h to allow viability detection. The MIC was determined as the lowest antimicrobial concentration at which no red color appeared (Langfield et al., 2004).
Synergy interaction assay
The checkerboard method (Pillai et al., 2005; Si et al., 2008; Aliskan et al., 2009) was performed to assess synergistic interactions between the antibiotics and natural antimicrobials as determined by the microbiological identification system VITEK® 2. Fractional inhibitory concentration (FIC) index was calculated for the natural antimicrobials (drug A) along with the antibiotics (drug B) to which the bacteria were normally resistant (Pei et al., 2009). The concentrations of the agents used were serially diluted two-fold from their MIC value. The formulas used are listed below (Pillai et al., 2005; Santiesteban-Lopez et al., 2007):
An FIC index ≤0.5 suggests a synergistic interaction; an FIC index >0.5 but ≤4.0 indicates “no interaction”; an FIC >4.0 reveals antagonism (Zhao et al., 2002; Odds, 2003; Hemaiswarya et al., 2008).
Investigation of physiological aspects between bacteria treated and untreated by natural antimicrobials
Six milliliters of stationary phase cultures, with and without 0.625 mM/mL cinnamaldehyde treatment, were collected and washed three times with PBS (Sigma-Aldrich). The pellets were resuspended with 4 mL of extract buffer (0.1 M phosphate buffer, pH 7, containing 2% ascorbate, and 0.1% 2-mercaptalethanal; Sigma-Aldrich). The cell densities were adjusted roughly equal by plating and growing in TSB into stationary phase to achieve 108–109 CFU/mL and serially diluted to the appropriate concentration. The 4 mL of bacterial suspension was concentrated and sonicated for 1 min, boiled for 10 min, quickly cooled on ice, and centrifuged at 4°C to remove cell debris. The supernatant was stored at −20°C until use. After filtration to remove precipitated proteins, 4 μL of the clear supernatant was injected into an Aquasil C18 column, 150×4.6 mm, 3 μm (Thermo Electron Corporation) on a UPLC system (Agilent 1100) equipped with an ultraviolet detector (range at 260–280 nm). The mobile phase was 99:1 of acetonitrile-phosphate buffer under linear gradient elution conditions. The flow rate was 0.4 mL/min. Differences among the identical peaks from different samples were compared by using Microsoft excel®.
Results and Discussion
Bacterial isolates from feed
A total of 20 isolates were selected from feed samples. After 3–4 subcultures, there were 17 successful isolates left, 5 of which were from duck feed, 9 from cow feed, and 3 from hog feed. All isolates were found to be Gram-negative bacteria, possibly as a result of the enumeration agar for all isolates (TSA), which is ideal for Gram-negative bacterial growth. Six selected isolates were then evaluated by the VITEK® 2 microbiological identification system (Table 1). The Moraxella spp. was found in both duck and hog feed and Rhizobium radiobacter was found in both cow and hog feed. The remaining two samples identified were K. oxytoca and S. paucimobilis from duck feed sample #5 and cow feed sample #5. Among 15 species in the Moraxella genus, some of which show opportunistic infection capability, Moraxella catarrhalis is clinically the most important species (Marshall and Jones, 1999). R. radiobacter has been recognized as an opportunistic human pathogen as most patients with R. radiobacter infection have debilitating underlying diseases (Lai et al., 2004), whereas K. oxytoca is an opportunistic pathogen that can cause colitis in humans (Stock and Wiedemann, 2001). Although S. paucimobilis is a microorganism of low clinical virulence, it can cause infections, which can lead to septic shock in healthy as well as immuno-compromised individuals (Lin et al., 2010).
Natural antimicrobial susceptibility testing
Natural antimicrobials, namely, eugenol, carvacrol, thymol, and cinnamaldehyde, at various concentrations, 10, 5, 2.5, 1.25, and 0.625 mM/mL, were investigated by testing their efficacy on selected bacterial animal feed isolates (Table 2). Higher concentrations (10 and 5 mM/mL) inhibited all bacterial growth, whereas 2.5 mM/mL produced the largest zones of inhibition of the remaining concentrations tested. Among the four different antimicrobials, carvacrol and eugenol showed better inhibitory effects, with larger inhibitory zones, compared with the remaining two compounds. This is consistent with the results by Friedman et al. (2002), who reported the descending antimicrobial order of carvacrol>cinnamaldehyde>thymol in a microplate assay against E. coli. Ethanol alone was found to have no effect on the growth of the bacteria.
The designation “−” indicates that there was no bacterial growth.
The designation “+” indicates that there was no noticeable zone of inhibition.
It has been reported that carvacrol, which is present in the essential oil fractions of oreganum (60%–70% carvacrol) and thyme (45% carvacrol), and its isomer thymol (with its hydroxyl group located at the meta position) are effective against E. coli (Dorman and Deans, 2000; Burt et al., 2005), L.monocytogenes (Singh et al., 2003), Shigella sonnei and Shigella flexneri (Bagamboula et al., 2004), and Bacillus cereus (Valero and Giner, 2005). Cinnamaldehyde and eugenol are major active components from cinnamon and clove essential oils and have shown promising antimicrobial capability against a number of food borne pathogens (Demo et al., 2001; Valero and Giner, 2005). To date, eugenol, cinnamaldehyde, thymol, and carvacrol have been registered by the European Commission and are considered to present no risk to human health (Burt, 2004); however, there has been a limited number of reports on their antimicrobial activity against K. oxytoca and S. paucimobilis.
Antibiotic resistance evaluation
Resistance patterns of the two tested isolates, alone and pretreated with the four natural antimicrobials with 0.1 mM/mL, were performed with the VITEK® 2 microbiological identification system. These results are shown in Table 3. Both duck #5 and cow #5 isolates, namely, K. oxytoca and S. paucimobilis (Table 1), displayed a variety of antibiotic resistance with different MIC values, among which, both bacteria showed great resistance to aztreonam, nitrofurantoin, and trimethoprim, whereas ciprofloxacin was the most effective antibiotic against both isolates. The MIC values of cefotetan, ceftazidime, ceftriaxone, ertapenem, and levofloxacin for cinnamaldehyde pretreatment of duck #5, K. oxytoca, were less than antibiotics alone, indicating an additive or synergistic effect, whereas the MIC values of ciprofloxacin increased, indicating an antagonistic effect with cinnamaldehyde. The rest of the available data from Table 3 remained the same MIC values between isolates alone and when pretreated with different natural antimicrobials.
The designation “–” indicates these combinations were not tested.
MIC, minimal inhibitory concentration.
Interestingly, current observations were the opposite to those reported by Stock and Wiedemann's (2001), who found that K. oxytoca was sensitive to aztreonam, trimethoprim/sulphamethoxazole, and nitrofuratoin, with MIC values of more than 16 mM/mL and intermediately sensitive to ampicillin, whereas the MIC value in the present study was 2 mM/mL. The MIC values of the listed antibiotics against S. paucimobilis are comparable with the results for K. oxytoca, with several exceptions, including cefotetan, ceftazidime, ceftriaxone, cefepime, and levofloxacin, with different MIC values. Both bacteria were challenged with four natural antimicrobials, which interfered with phenotypic expression of genetically determined antibiotic resistance.
MIC determination
The MIC was defined as the lowest antimicrobial concentration that prevented visible bacterial growth (Santiesteban-Lopez et al., 2007; O'Bryan et al., 2008; Li et al., 2010). The MIC results for four natural antimicrobials against two bacterial strains are presented in Table 4. S. paucimobilis showed 2–15-fold lower MIC values for the four natural antimicrobials when compared with K. oxytoca. Among these four natural antimicrobials, cinnamaldehyde and eugenol shared the lowest MIC values of 0.156 mM/mL against the S. paucimobilis cow #5 isolate. Cinnamaldehyde and carvacrol showed the highest MIC values of 2500 mM/mL against the K. oxytoca, duck #5 isolate.
Pei et al. (2009) found that E. coli was inhibited by eugenol, cinnamaldehyde, thymol, and carvacrol, at 9.7, 2.6, 3, and 2.6 mM/mL, respectively. In the present study (Table 4), the MIC values of both bacteria against the same four natural antimicrobials showed completely different results except for K. oxytoca, which showed similar results with cinnamaldehyde and carvacrol. The much lower MIC values of the rest of the results indicated that both bacteria are much more sensitive when compared with the antibiotic-resistant E. coli in Pei et al.'s (2009) study. However, the present results for K. oxytoca are consistent with Helander et al. (1998), who showed that carvacrol, thymol, and cinnamaldehyde inhibited E. coli and S. Typhimurium at 1 to 3 mM/mL. This also suggests that S. paucimobilis is more sensitive compared with K. oxytoca against these four natural antimicrobials.
Synergistic interaction study
According to the formulas mentioned above (Pillai et al., 2005; Santiesteban-Lopez et al., 2007), the FIC index values are listed in Table 5. For S. paucimobili, a synergistic interaction was found for both ampicillin and nitrofurantoin in all tested natural antimicrobials, except that there were three “no interactions” for cinnamaldehyde and ampicillin, carvacrol and nitrofurantoin, and Eugenol and ampicillin combinations with FIC index values of 1, 1, and 0.625, respectively. The FIC index values for K. oxytoca were generally higher, with only two “synergistic interactions,” namely, carvacrol and ampicillin, and carvacrol and nitrofurantoin combinaions, with FIC index values of 0.375 and 0.15, respectively, which indicated that the natural antimicrobials synergistically enhanced the efficacy of the antibiotics to which these bacteria were normally resistant (Odds, 2003; Santiesteban-Lopez et al., 2007; Si et al., 2008).
Synergy=an FIC index ≤0.5. FIC index >0.5 and ≤4.0 indicates “no interaction.” An FIC >4.0 indicates antagonism between the two agents.
FIC, fractional inhibitory concentration.
When antibiotics and natural antimicrobials were tested in paired combinations for possible synergistic effects against S. paucimobilis and K. oxytoca, a considerable reduction in the MICs of the antibiotics was noted (Tables 3 and 5). Synergy is generally recognized when the combined effect of two compounds is greater than the sum of the effects of each compound alone (Odds, 2003; Kobilinsky et al., 2007). Among four natural antimicrobials, thymol and carvacrol were found to be very effective (FIC ≤0.5) in combination with all antibiotics tested against S. paucimobilis and K. oxytoca, respectively; Eugenol or cinnamaldehyde was less effective in reducing resistance to the antibiotics, though they had the same effect as thymol and carvacrol when used alone. The mechanisms behind this phenomenon, the reduction in antibiotic resistance by natural antimicriboals, are unknown; however, it is possibly due to certain structural changes in bacteria cells. Some accepted mechanisms of antimicrobial synergistic interactions are the sequential inhibition of a common biochemical pathway, inhibition of protective enzymes, combinations of cell wall-active agents, and use of cell wall-active agents to enhance the uptake of other antimicrobials (Eliopoulos and Moellering, 1991).
The natural antimicrobials may facilitate penetration of the antibiotics through the outer layers of the bacterial cell wall, act by blocking the inhibitory effects of protective enzymes, or interfere with single or multiple metabolic targets of the antibiotic (Hemaiswarya et al., 2008). Pei et al. (2009) hypothesized that the hydroxyl group on eugenol may combine with proteins, inhibiting enzyme action; the carbonyl group on cinnamaldehyde may adhere to proteins to prevent the action of amino acid decarboxylases. Carvacrol acts as a transmembrane carrier of monovalent cations by exchanging its hydroxyl proton for another ion (Ultee et al., 2002). Accumulated carvacrol occupies more than the normal amount of space between fatty acid chains, resulting in conformational changes of the phospholipid bilayer. A disturbance of the van der Waals interactions between the lipid acyl chains in the membrane causes fluidization of membrane lipids (Ultee et al., 1999). Similar to carvacrol, thymol was hydrophobic and prone to disturb the outer membrane of Gram-negative bacteria, releasing lipopolysaccharides, and increasing the permeability of the cytoplasmic membrane to ATP (Helander et al., 1998; Lambert et al., 2001).
Physiological differences of bacteria isolates between natural antimicrobial-treated and -untreated cells
Cow isolate #5, S. paucimobilis, and duck isolate #5, K.oxytoca, together with L. monocytogenes ATCC 19115 and E. coli isolates, alone and treated with four natural antimicrobials, thymol, cinnamaldehyde, eugenol, and carvacrol, were investigated by UPLC system (Agilent 1100) and 20 graphs were produced (data not shown). There were no systematic difference between bacteria alone and natural antimicrobial treatments, except that two identical peaks appeared with bacteria alone (i.e., L. monocytogenes ATCC 19115 and E. coli) but disappeared with cinnamaldehyde treatments (Fig. 1). The two peaks appeared at retention times of 2.66 and 3.35 with cow #5, S. paucimobilis, and 2.67 and 3.35 with duck #5, K. oxytoca. The two peaks are suspected to be two identical metabolic products produced by both of the isolates without cinnamaldehyde treatments. Further research in this area is needed and the literature related to the metabolism of cinnamaldehyde-treated cells is limited. Niu et al. (2006) reported that cinnamaldehyde would influence signaling pathways of quorum sensing by interfering with the binding of 3-hydroxy-C4, 3-oxo-C6-HSL and AI-2 synthetic pathway. Brackman et al. (2008) discovered that cinnamaldehyde interferes with AI-2 based quorum sensing by decreasing the DNA-binding ability of LuxR to bind to its target promoter sequence. Cinnamaldehyde not only affected the bacterial production of multiple virulence factors but also reduced the mortality of food products exposed to bacteria. Thus, the two metabolites that disappeared may be related to quorum-sensing pathways.
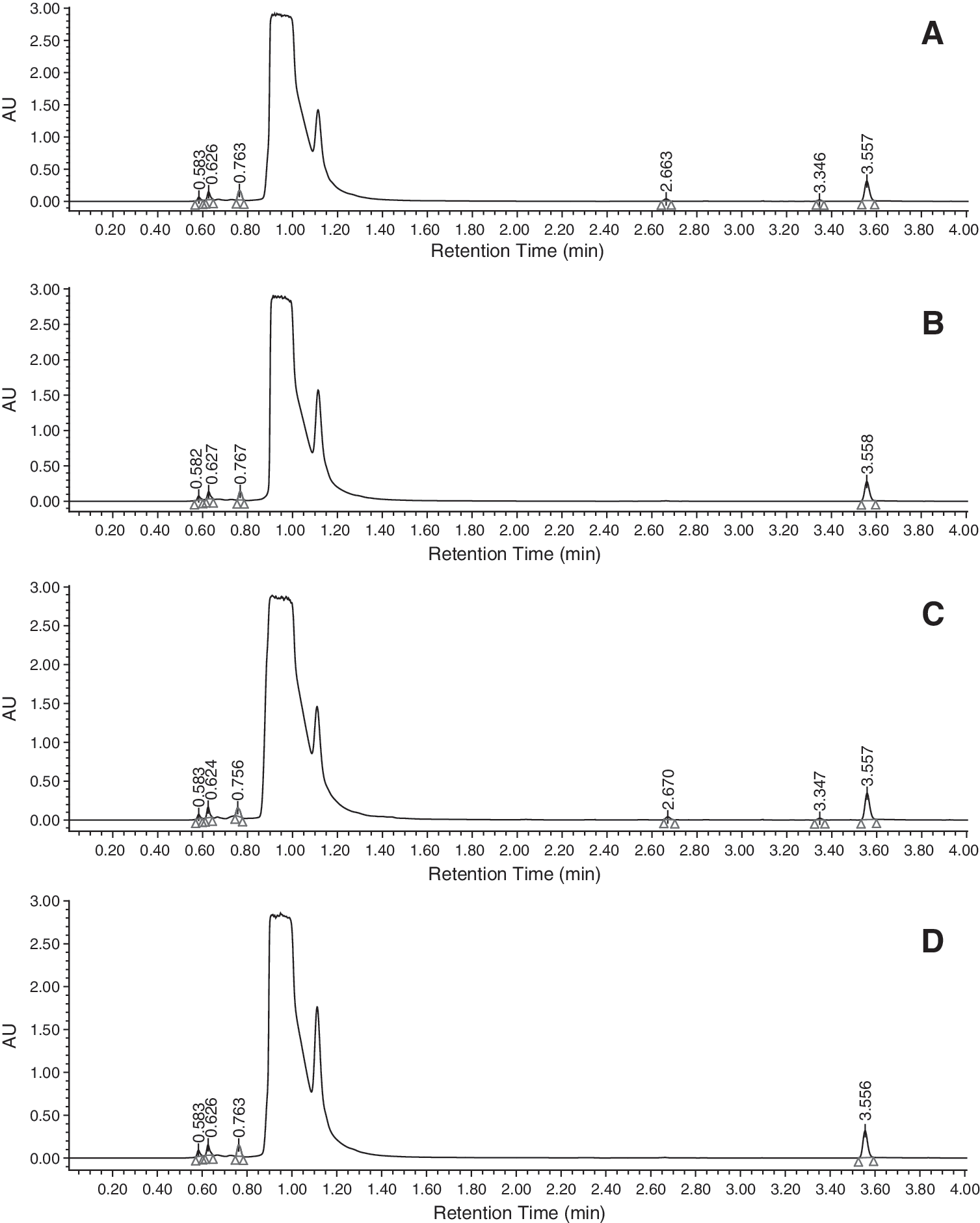
Physiological differences of bacteria isolates between natural antimicrobial-treated and -untreated cells.
Conclusions
While new antibiotic drug development is both difficult and expensive, new methods to reduce the development of antibiotic resistance by pathogenic organisms in food are needed. The present work suggested that substituted phenolic compounds, eugenol, cinnamaldehyde, thymol, and carvacrol, have the potential for use to control the development of antibiotic-resistant bacteria in animal feed. The substituted phenolic compounds examined were effective inhibitors of the antibiotic-resistant bacteria studied when used individually. The synergistic interaction results showed that the natural antimicrobials, specifically thymol and carvacrol, were able to substantially lower the level of antibiotic resistance in S. paucimobilis and K. oxytoca, respectively. Further, physiological studies estimated that two identical metabolites that disappeared with cinnamaldehyde treatments might be associated with quorum-sensing pathways.
Footnotes
Acknowledgments
The present work is supported by Shanghai Academy of Agricultural Sciences Start-up fund. We are grateful to Dr. Joshua Gurtler, Food Safety Intervention Technologies, ARS Eastern Regional Research Center, U.S. Department of Agriculture, for his kind proofreading.
Disclosure Statement
No competing financial interests exist.