
Abstract
To examine the serotype specificity of inlH, which encodes the virulence-associated surface protein InlH related to the intracellular survival of Listeria monocytogenes in mice, the presence of inlH in 337 L. monocytogenes strains, representing 11 different serotypes, was studied. A total of 106 strains representing 3 serotypes and 14 pulsed-field gel electrophoresis (PFGE) types were positive for inlH by polymerase chain reaction. inlH was present in all 99 serotype 1/2c and 3 serotype 3c strains. Moreover, 4 out of 129 (3%) serotype 1/2a strains carried inlH. All 106 strains representing serotypes 1/2b, 3a, 3b, 4a, 4b, 4c, 4d, and 7 and 125 out of 129 (97%) serotype 1/2a strains were inlH-negative. The coding sequences of the inlH genes of eight L. monocytogenes strains representing three serotypes and five PFGE types were identical. These results suggest that inlH is specifically present in serotype 1/2c, 3c, and a small fraction of 1/2a L. monocytogenes strains and exists as a single allele.
Introduction
S
The number and type of internalin genes differ among L. monocytogenes strains (Nelson et al., 2004). While inlA, inlB, and inlC are present in nearly all L. monocytogenes strains (Doumith et al., 2004; Tsai et al., 2006; Chen et al. 2009), inlF is present only in majority of serotype 1/2c and 3c and some serotype 1/2a and 3a strains (Chen et al. 2009). Most serotype 1/2c and 3c strains and half of serotype 1/2a and 3a strains have Lmo2026 (Chen et al., 2009). The prevalence of inlH varies among studies (Raffelsbauer et al., 1998; Doumith et al., 2004; Tsai et al., 2006; Chen et al., 2009). To examine whether inlH is serotype specific, we studied the presence of inlH in 337 L. monocytogenes strains by polymerase chain reaction (PCR).
Materials and Methods
Bacterial strains
A total of 326 L. monocytogenes strains from our culture collection, serotyped with commercial antisera (Denka-Seiken, Tokyo, Japan), and 11 L. monocytogenes reference strains were studied (Tables 1 and 2).
All human isolates originate from clinical cases, and animal isolates originate from clinical cases or from healthy carriers.
Polymerase chain reaction
Based on published DNA sequences of inlH of serotype 1/2a L. monocytogenes strains and inlD of serotype 1/2a, 1/2b, 3b, and 4b strains, one pair of PCR primers was designed for inlH, and two pairs for inlD (Table 3). Strains negative with serotype 1/2a-specific inlD primers were further examined with inlD primers specific for serotypes 1/2b, 3b, and 4b. In short, 50-μl PCR mixtures contained 1 μl of heated cell lysate as template, 0.4–0.5 μM of each primer (Oligomer, Helsinki, Finland), 0.2 mM of dNTP (10 mM dNTP Mix; Finnzymes, Espoo, Finland), and 2 U of DyNAzyme II DNA Polymerase (Finnzymes) in 1× reaction buffer. The PCR products were visualized in agarose gel.
Primers are presented in 5′ to 3′ direction.
Primer specific for serotype 1/2a.
Primer specific for serotypes 1/2b, 3b, and 4b.
From inlH-positive strains, a 270-bp fragment was amplified. From some inlH-negative strains, approximately 2100-bp fragments were amplified from the consecutive genes inlC2 and inlD.
From inlH-positive strains, a 794-bp fragment was amplified. From some inlH-negative strains, approximately 2706-bp fragments were amplified from the consecutive genes inlC2 and inlD.
PCR, polymerase chain reaction.
Pulsed-field gel electrophoresis
In situ DNA isolation and pulsed-field gel electrophoresis (PFGE) of inlH-positive strains were performed according to Autio et al. (2002). AscI macrorestriction patterns were analyzed, using Bionumerics software version 4.5 (Applied Maths, Kortrijk, Belgium).
inlH sequencing
From eight L. monocytogenes strains (AL4E, RE2E, HE19E/1, LL28/1, RT24E, HL35E/1, LM120/1, and LMH18), an approximately 1970-bp fragment containing the entire coding sequence of inlH was amplified by PCR and sequenced using primers presented in Table 3. Bionumerics software version 4.5 was used for assembly.
Results and Discussion
inlH was detected in 106 (31%) L. monocytogenes strains from at least five European countries, including all 99 serotype 1/2c strains, all three 3c strains, and four (3%) 1/2a strains. A total of 125 (97%) serotype 1/2a strains and all 106 strains representing serotypes 1/2b, 3a, 3b, 4a, 4b, 4c, 4d, and 7 were inlH negative. Our results are supported by Chen et al. (2009) reporting 14 out of 17 serotype 1/2c or 3c strains and some 1/2a strains from Chinese food systems to carry inlH, and Tsai et al. (2006) also reported that rare 1/2a strains from New York State carry inlH. Both studies showed absence of inlH in serotypes 1/2b, 3b, 4a, and 4b.
Doumith et al. (2004) reported inlH in serotype 1/2c and 3c strains, but also in strains representing serotypes 1/2a, 1/2b, 3a, 3b, 4b, 4c, 4d, 4e, and 7. As suggested by Jia et al. (2007), the array-based result by Doumith et al. (2004) may have been due to false-positive hybridization of the inlH probe to inlC2, which is highly similar to inlH. The similarity of inlH and inlC2 derives from the emergence of the mosaic inlH as a fusion of the consecutive inlC2 and inlD, caused by deletions in the 3′end of inlC2 and the 5′end of inlD, in an ancestral L. monocytogenes (Raffelsbauer et al., 1998). Moreover, Raffelsbauer et al. (1998) found 11 L. monocytogenes strains representing serotypes 1/2a, 1/2b, 1/2c, 3a, 3b, 3c, 4a, 4b, and 4e positive for inlH with PCR, whereas only 4c and 4d strains were inlH negative. Thus, to accurately assess the differences between results by Raffelsbauer et al. (1998) and those from other studies (Tsai et al., 2006; Chen et al., 2009), we examined the presence of inlH in 11 reference strains (Table 2) with primers used by Raffelsbauer et al. (1998). Using PCR mixture and reaction conditions presented in Materials and Methods and Table 3, respectively, only EGDe and ATCC 7644, the only inlH-positive reference strains with our primers, showed a 794-bp fragment specific for inlH, supporting our findings. We do not know the reason for the discrepancy between our results and those by Raffelsbauer et al. (1998), but may explain it by differences in PCR performance. Another explanation is different selection of strains; however, this is opposed by the fact that the aforementioned studies with strains from even two other continents reported results similar to ours (Tsai et al., 2006; Chen et al., 2009).
To complement our inlH PCR result, we also studied the presence of inlD. All 106 inlH-positive strains were negative for inlD and all 231 inlH-negative strains, except the only serotype 4a strain, had inlD (Tables 1 and 2). Similarly, inlH-positive L. monocytogenes strains in studies by Tsai et al. (2006) and Chen et al. (2009) were inlD-negative, whereas all inlH-negative strains, except two serotype 4a strains, were positive for inlD. Based on all three studies with European, American, and Asian strains, and international reference strains, inlH appears to be present solely in L. monocytogenes serotypes 1/2c and 3c and a few strains of serotype 1/2a.
In addition to the closely related serotypes 1/2c and 3c, inlH was present in 3% of serotype 1/2a strains. The presence of inlH in three different serotypes likely derives from a common ancestor of all inlH-positive strains. A phylogram of 19 inlC2 sequences and the 5′ end of six inlH sequences publicly available (Fig. 1) suggests that inlH emerged in an ancestral serotype 1/2a strain. Further genetic events in the progeny of this first inlH-positive strain may have yielded serotypes 1/2c and 3c. Results by Ward et al. (2004) support our hypothesis about serotypes 1/2c and 3c emerging from serotype 1/2a strains.

Maximum likelihood phylogenetic tree for corresponding 1000-bp sequences from 5′end of inlC2 or inlH of Listeria monocytogenes strains representing serotypes 1/2a, 1/2b, 1/2c, 3b, 4a, and 4b, available in public databases.
PFGE typing of all 106 inlH-positive strains yielded 14 pulsotypes (Fig. 2). Eight strains representing three serotypes and five pulsotypes carried an identical 1647-bp inlH allele (EMBL Nucleotide Sequence Database, accession numbers AM905839-AM905846). They were also identical to all six inlH sequences published (Raffelsbauer et al., 1998; Glaser et al., 2001; Tsai et al., 2006; Broad Institute). inlH apparently occurs with one allele worldwide.
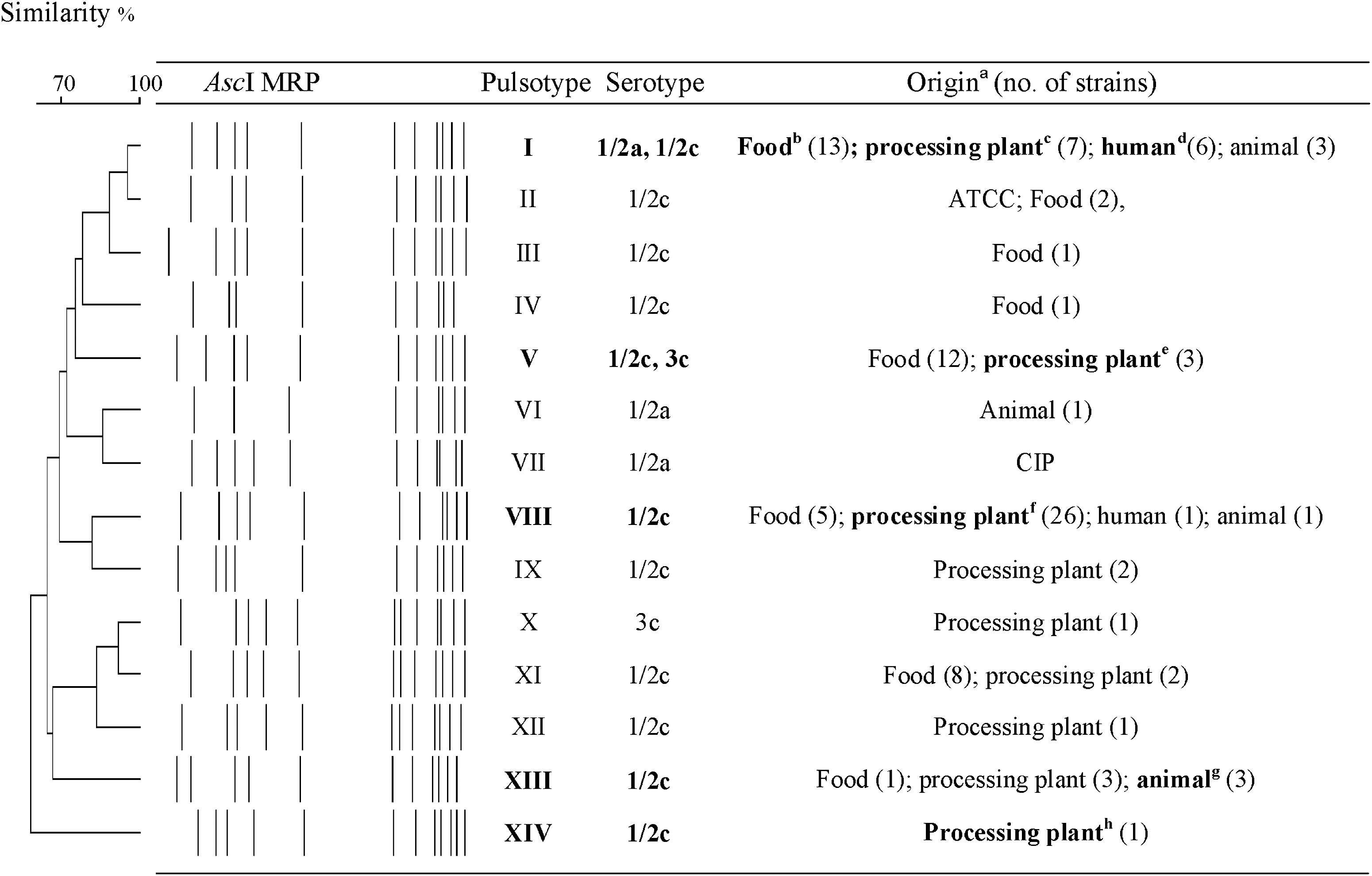
Dendrogram of AscI macrorestriction patterns (MRPs) represented by inlH-positive L. monocytogenes strains. Similarity analysis of MRPs was performed, using the Dice coefficient, and clustering was performed by unweighted pair group method using arithmetic averages (UPGMA) (position tolerance 1.0). The cophenetic correlation of the dendrogram is 58%. The strains for which inlH was sequenced are indicated in boldface. aATCC, American type culture collection, source of strain ATCC 7644; CIP, The collection of Institut Pasteur, source of strain EGDe. bOrigin of strain LM120/1. cOrigin of strain LMH18. dOrigin of strain AL4E. eOrigin of strains HE19E/1 and HL35E/1. fOrigin of strain RT24E. gOrigin of strain LL28/1. hOrigin of strain RE2E.
In conclusion, all L. monocytogenes serotype 1/2c and 3c strains tested had inlH. Few serotype 1/2a strains were inlH positive, whereas inlH was not detected in any serotype 1/2b, 3a, 3b, 4a, 4b, 4c, 4d, and 7 strains. All inlH-negative strains carried inlD, except the only serotype 4a strain studied, which lacked both genes. L. monocytogenes strains representing different sero- and pulsotypes carry an identical inlH gene. To our knowledge, inlH is the first virulence-associated gene present mainly in serotype 1/2c and 3c L. monocytogenes strains.
Footnotes
Acknowledgments
This research was carried out at the Finnish Center of Excellence in Microbial Food Safety Research. The study was supported by the Walter Ehrström Foundation.
Disclosure Statement
No competing financial interests exist.