
Abstract
Objectives
Materials and Methods
Results
Discussion and Conclusion
Introduction
S
Salmonella infections in animals and humans mainly spread via the oral–fecal route. In most cases, humans get infected by consumption of contaminated and insufficiently cooked food. Well-known sources for Salmonella Enteritidis are eggs, poultry meat, and related products (Wray and Wray, 2000; EFSA, 2010). Control measures in the whole food chain, from farm to fork, are needed to decrease the prevalence of salmonellosis in humans (Davies and Wray, 1996; Hald et al., 2004; Voetsch et al., 2004; Forshell and Wierup, 2006; Collard et al., 2007; Gast, 2007; Singer et al., 2007; Bollaerts et al., 2008, 2009; Bertrand et al., 2010). On top of the sanitary consequences, the disease constitutes an economic burden due to an increased medical and sometimes hospital cost (the annual cost of Salmonella in the United States in 2009 was estimated at USD 2,649,413,401) (ERS USDA, 2010), as well as loss of productivity of diseased people and damage to the poultry or pork market.
After a continuous increase of Salmonella spp. infections in the Belgian human population from 6092 in 1986 to 15,774 isolates registered in 1999, cases began to decrease. In 2007, 3975 isolates were registered, of which 987 were Salmonella Enteritidis (25%) (ISP-WIV, 2008). The tightened sanitary measures in the poultry breeders and layers as well as vaccination against Salmonella Enteritidis that were implemented since 2004 probably have contributed to the decrease of human cases of Salmonella Enteritidis (Voetsch et al., 2004; Forshell and Wierup, 2006; Collard et al., 2007).
Identifying the different Salmonella serotypes and phage types (PTs) from human and animal sources is of major importance for tracing back the source of infection after or during outbreaks, and in turn for implementing prevention and control measures (Altekruse et al., 1993; Gast and Beard, 1993; van Duijkeren et al., 2002; Hald et al., 2004; Gast, 2007; Singer et al., 2007; Voetsch et al., 2009).
Based on European legislation 92/117/EEC and 2160/2003/EC, Belgium organized official sampling of poultry breeding flocks from 1993 onward and published national legislation describing the surveillance programs (Royal Decrees of 22 May 2005, 2005022397 and 27 April 2007, 2007022784). In 2004, the European Union (EU) decision 2004/665/EC imposed a coordinated monitoring program of Salmonella among layers following the technical specifications laid down in DG SANCO/34/2004. The aim of this coordinated monitoring program was to set up target prevalence to be reached by each Member State in the following years.
Based on the results of this EU-coordinated monitoring program, the PT of all isolated Salmonella Enteritidis collected in Belgium in 2005 were compared with the Salmonella Enteritidis PT isolated from humans. The aim of the present study was twofold. First, the total PT distribution over the whole year 2005 in both populations was investigated to determine whether they were statically comparable and, second, whether the monthly distribution of the most common PT of Salmonella spp. differed significantly in both populations.
Materials and Methods
Data collection
Coordinated EU monitoring program in poultry layers
During the year 2004–2005, a national survey (European coordinated monitoring) was organized in accordance with the Commission Decision 2004/665/EC to estimate the prevalence of Salmonella in each member state and to define a target prevalence of Salmonella in whole Europe for the poultry laying sector. For Belgium, according to EFSA's technical specifications, 1043 samples from 149 layer flocks in different holdings were taken during the year 2005, from February until September. A total of seven samples, five from fecal material and two environmental samples, were taken per flock in each holding.
Samples were sent to the two regional laboratories, “Association Regionale de Santé et d'Identification Animales” (ARSIA) and “Dierengezondheidszorg Vlaanderen” (DGZ), for Salmonella isolation and detection according to ISO6579:2002/Amd 1:2007. In case of isolation of Salmonella, one isolate per sample was serotyped following the Kauffmann-White scheme (Grimont and Weill, 2007) at the National Reference Laboratory (NRL) for Salmonella animal health (CODA-CERVA). All Salmonella Enteritidis isolates were phage typed at the National Reference Centre for Salmonella and Shigella (NRCSS, Public Health Institute) according to the recommendations of the Health Protection Agency Service (Colindale, United Kingdom) (Threlfall and Frost, 1990).
Human data
Belgian Salmonella isolates collected from human gastroenteritis reported cases by several peripheral clinical laboratories were transferred to the NRCSS for Kauffmann-White serotyping (Grimont and Weill, 2007). All the human cases of Salmonella Enteritidis reported and submitted for phage typing during the year 2005 at the NRCSS were used in this study.
Descriptive statistics
Results of the Salmonella prevalence survey carried out in layers holdings, and results of the NRCSS annual report of human outbreaks were examined and analyzed descriptively to obtain insight on the prevalence of Salmonella, the different serotypes, and Salmonella Enteritidis PT's monthly and yearly distribution (SAS 9.2).
The distribution of the Salmonella Enteritidis human cases and the sampled layer holdings were geographically represented in Arc-GIS 9.3 (ESRI BeLux SA, Wemmel, Belgium). For geo-localization, the patient's home postal code was used, if available, or alternatively, the postal code of the clinical lab where the sample was analyzed. A separate map was created to represent the two most frequent isolated Salmonella Enteritidis PT in humans and layers.
Finally, a time cluster analysis was carried out using the Scan Test statistic described by Carpenter and Ward (2003) to determine whether the clusters of PT observed monthly in both populations, respectively, were significant or not, based on the exact approximation of the p-value.
Data analysis
To investigate whether a possible common trend existed between the different Salmonella Enteritidis PT isolated in humans and layers, different statistical tests were used.
Due to the scarce data and the non-normality distribution in some of PT distributions over the whole year, parametric and non-parametric tests were chosen to investigate the difference in each PT distribution among humans and layers. First, the distribution of the total different PT isolated in both layer and human populations during the whole year 2005 was investigated and compared using a Kruskal–Wallis (KW) test (StatXact® 4.0.1.) for K independent samples. The class variables were the different PT proportions and the dependent variables were human and layers. Due to an unbalanced design in the datasets (474 PT results in humans and 143 in layers), the relative proportion of a given PT in comparison to all other PTs in that population (human/layers) that year was used.
The two most frequent PT (PT21 and PT4) were studied further to determine whether their monthly distribution was different in both populations. Non-parametric Wilcoxon Mann–Whitney (MW) test was used to compare the monthly proportions of the same PT in the human and layer population. Only the months where counts in human and layers were present were investigated (February to September). The results were comforted by the parametric test analysis of variance test (SAS 9.2). The class variables were PT21 and PT4 in layers and the dependent variables were, respectively, PT21 and PT4 in humans. Due to the unbalanced design, the generalized linear model (GLM) was used instead to run the analysis of variance test.
Results
Descriptive statistics
Thirty-eight percent (95% CI: 32.1–44.4) of the poultry holdings sampled (flocks) were Salmonella positive (i.e., at least 1 out of the 7 samples taken were positive for Salmonella spp.) and 27% (95% CI: 21.9–33.1) of these holdings were Salmonella Enteritidis positive. Twenty percent of the samples were positive for Salmonella and 13% for Salmonella Enteritidis. The within-holding Salmonella spp. prevalence ranged from 0% to 100%, but never reached 100% for Salmonella Enteritidis.
The total number of recorded Salmonella spp. human infections in Belgium in 2005 was 4916, out of which 2226 Salmonella Enteritidis were isolated. In the same year, 474 randomly selected Salmonella Enteritidis isolates were phage typed. Eight Salmonella outbreaks were reported for the year 2005. Only one case could be traced back and originated from tiramisu that was consumed by the patient.
Table 1 illustrates the distribution of the different Salmonella serotypes in the human and layer population in 2005.
In both populations it is clear that the most predominant serotype was Enteritidis, followed by Mbandaka, Braenderup, and Livingstone in layers. In humans, Typhimurium was the second most frequent isolated serotype, followed by Brandenburg, Derby, Virchow, and Infantis.
As for Salmonella Enteritidis, the PT commonly found in both populations were PT1, 4, 6, 6a, 8, 14b, 21, 35, and RDNC 69. PT21 followed by PT4 were the most frequently isolated PTs from both populations. Some PTs were exclusively found in humans (1a, 2, 7a, 9a, 11, 12, 13, 23, 31, 34, 37, RDNC/P14b, RDNC/P20a, RDNC/P21a, RDNC/P3, RDNC/P30, RDNC/P5a, RDNC12, RDNC214, RDNC27, RDNC39, RDNC804, and RDNC811), whereas others were only found in layers (3, 4a, 6a/35, 7, 10, 35W, 38).
Figures 1 and 2 map the distribution of Salmonella Enteritidis cases in human and layer populations.
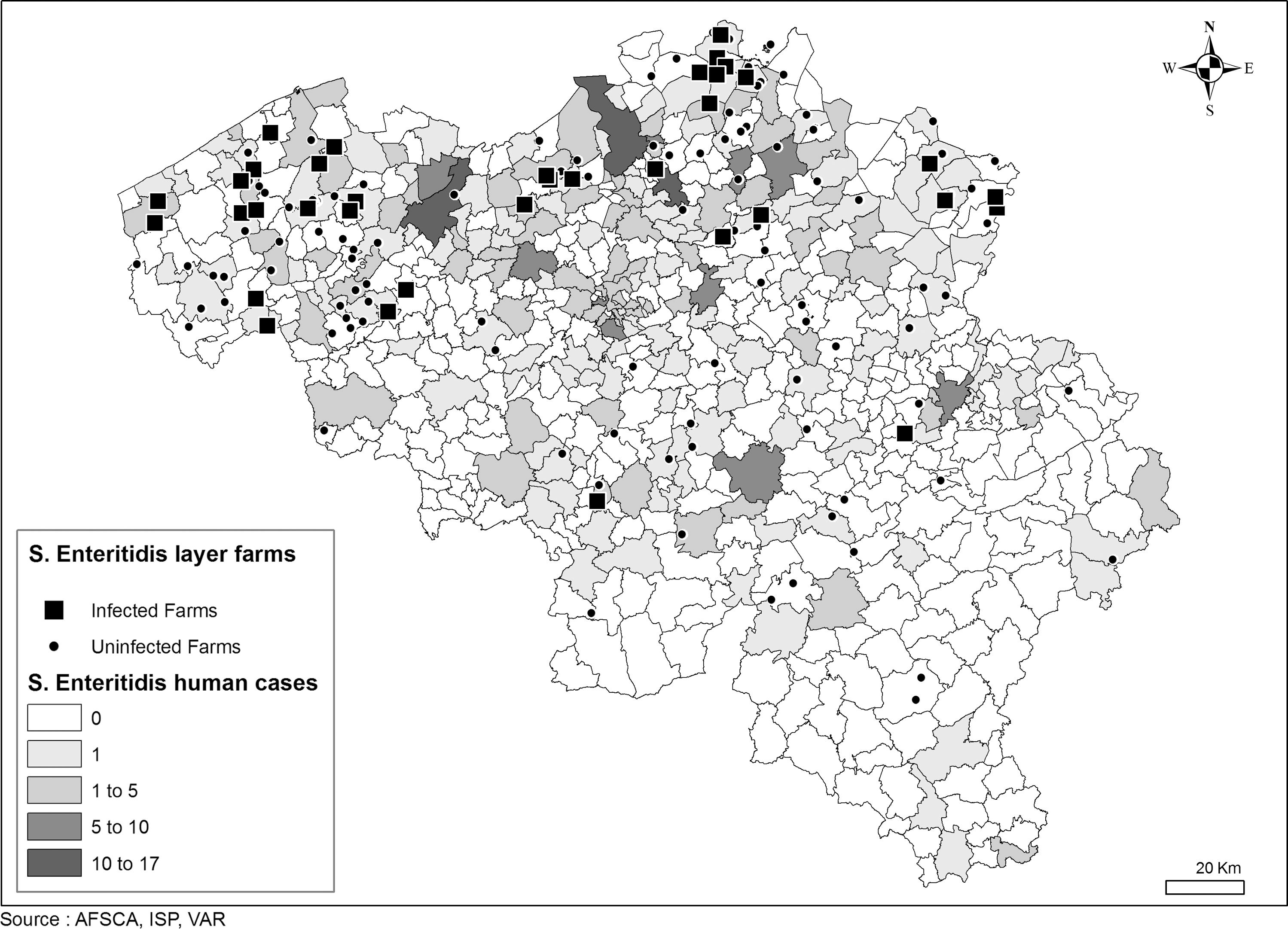
Distribution of Salmonella Enteritidis cases in human and layers, as well as the sampled layer holdings.
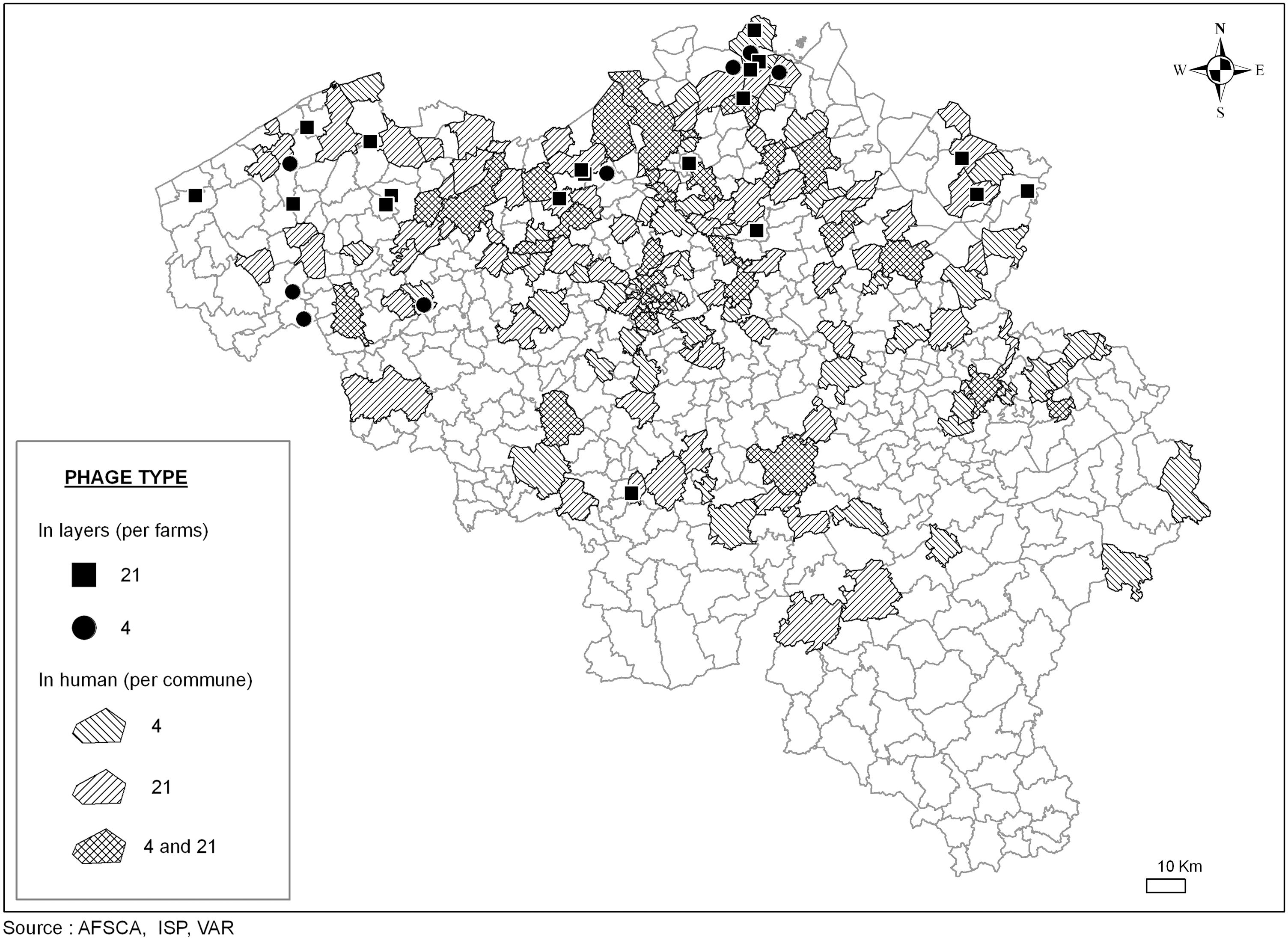
Distribution of Salmonella Enteritidis PT4 and PT21 in human and layer population. PTs, phage types.
The resolution displayed in Figure 1 is at municipality level. The higher number of holdings sampled in the northwest of the country is related to the higher density of layer holdings in that part of the country. The highest number of Salmonella Enteritidis human cases (more than 10/municipality) were found in the provinces of Oost-Vlaanderen and Antwerpen. The Salmonella Enteritidis PT distributions in both populations are clustered in the north of the country (Flanders region). There seems to be no geographical correlation between the human PT distribution and those of layers.
Figure 3 displays the evolution of the monthly relative proportions of Salmonella Enteritidis PT4 and PT21 in in humans and layers over the year 2005. The relative proportion of the given PT in the Y-axis is the proportion of the given PT to the total of the respective PT (4 or 21) isolated in that month.

Monthly evolution of Salmonella Enteritidis PT4 and PT21 in humans and layers in 2005.
The time cluster analysis, conducted using the methods described by Carpenter and Ward (2003), identified significant clusters for all PT during the months May–June layers and May–July–August–September (p-values <0.05) for humans. Several simulations were done to find out the most significant time window (PT counts per month or per 2 month); a 2-month window frame was the most significant time width.
Separate time cluster analyses were done for PT4, PT21, and the other remaining PT together. PT21 was found to be the main PT responsible of these time clusters.
Data analysis
The KW test showed that the Salmonella Enteritidis PT distributions were only border line significantly different between humans and layers (p-value=0.0491).
PT21 followed by PT4 turned out to be the most frequent PT isolated in both populations. Therefore, the link between the two populations was investigated based on these two PT. The results of the univariate analysis showed that PT4 in layers (but not in humans) and PT21 in human and in layers followed a normal distribution. Therefore, parametric and non-parametric tests were applied together to estimate whether the monthly distribution of these two PT differed in both populations. Results illustrate no difference in monthly proportions of PT4 and PT21 in both populations (PT4 0.59 and 0.73, and PT21 0.31 and 0.35 for the p-values of the MW and the GLM test, respectively).
Discussion
Results of the present study enabled to demonstrate statically similar trends between human and layer populations for Salmonella spp. and for Salmonella Enteritidis PT monthly prevalence over the year 2005. PT21 was the most frequent isolated PT in Belgium during the year 2005; this was not the case in the other European member states where PT4 was the most frequently reported and isolated PT (Collard et al., 2007; EFSA, 2007). Also, the time cluster analysis (Fig. 3) revealed that the significant clusters of Salmonella cases in humans could be attributed to Salmonella in layers. PT21 was the main PT responsible for the clusters of cases in both populations, and a slight delay (humans after layers) was observed, which comforts this hypothesis. Also, the 2-month time cluster supports the hypothesis that these Salmonella epidemics could result from a transmission episode and not from random infection points (Carpenter and Ward, 2003). Furthermore, the nonsignificant difference found from the KW, MW, and F statistic, between both populations, indicates that a common trend can be assumed between human and layers for the 2 most frequent PT4 and PT21, and this trend seems to be related.
One successful outbreak investigation identified Salmonella Enteritidis PT21 in tiramisu consumed by the infected patient. This observation confirms that consumption of contaminated eggs may lead to human infections in Belgium.
The prevalence figures might be underestimated in both populations. Results from the European coordinated monitoring program in the Belgian laying sector have been explored. The prevalence of Salmonella spp. and Salmonella Enteritidis positive holdings of 38% and 27%, respectively, were already published in the EFSA reports (EFSA, 2007). Nevertheless, whether these figures reflect the true prevalence is questionable. Previous studies have shown that the sampling strategy has an influence on the detection of Salmonella (Carrique-Mas et al., 2008; Van Hoorebeke et al., 2009). However, for the present study it was assumed that the results reflected the true prevalence in Belgium. A remarkable observation was that various Salmonella serotypes as well as various PTs of Salmonella Enteritidis can be found on the same farm. This probably reflects the multiple sources of contamination for a layer farm (Forshell and Wierup, 2006; EFSA, 2007).
In humans, underreporting could lead to underestimation of the true prevalence. Mead et al. (1999) and Voetsch et al. (2004) mentioned that underreporting of human cases of gastroenteritis is because only clinical isolates are reported, only a small part of human clinical cases are sampled and analyzed, and that there are probably more subclinical Salmonella infections (Wray and Wray, 2000).
The cluster of Salmonella-positive layer holdings in the northern part of Belgium must be interpreted cautiously due to the higher density of poultry holdings in that region (Fig. 1). Similarly, the more abundant salmonellosis cases in humans may be related to the higher density of the habitants in the northern part of the country. After correction (n cases/N population) relative figures were comparable in the three regions (Brussels: 41/1,006,749=0.00004; Flanders: 318/6,043,161=0.00005; Wallonia: 100/3,413,978=0.00003). Although the main cause of foodborne Salmonella Enteritidis is considered to be eggs (Hald et al., 2004; de Jong and Ekdahl, 2006; Voetch et al., 2009; EFSA, 2010), it is difficult in this study to draw sound conclusions of human cases related to the spatial neighboring cluster of positive layer holdings. Table eggs are distributed to various points of sale, not necessarily close to the layer farm. Table eggs are also traded (import/export) or may come from backyard hens (not sampled in this study). Without these data, prevalence figures in relation with human cases are difficult to interpret. Time clusters are more relevant in this case, as illustrated in the present study.
Phage typing has been recognized as an efficient screening method for tracing back infection, before conducting genotyping (Altekruse et al., 1993; Gast and Beard, 1993; van Duijkeren et al., 2002; Hald et al., 2004; de Jong and Ekdahl, 2006; Kafatos et al., 2009; Voetsch et al., 2009). Although similar trends could be identified, knowing the exact sources and route of spread would require further investigation. Vertical and horizontal transmission routes must be considered. Therefore, investigating the vertical transmission from parent's flocks is important. For the horizontal transmission routes, besides contaminated food and feed consumption, other sources of infection should be considered such as common reservoirs in the environment, animated or unanimated vectors, and contaminated sewage water as recognized in previous studies (van Duijkeren et al., 2002; Hald et al., 2004; de Jong and Ekdahl, 2006; Forshell and Wierup, 2006; Gast, 2007; Singer et al., 2007; Vestby et al., 2009; Voetsch et al., 2009; EFSA, 2010). The present study enabled the identification of interesting features about the common trends in both populations. In addition, this statistical approach could be a useful tool for policy makers to identify common trends and to tackle public health matters efficiently.
Footnotes
Disclosure Statement
No competing financial interests exist.