
Abstract
Background:
Bacillus cereus is increasingly recognized as one of the causative agents of food poisoning in different parts of the world.
Methods:
In this study, molecular characterization of B. cereus toxigenic strains isolated from food in Jordan was performed. A total of 202 food samples were collected from local markets and restaurants.
Results and Discussion:
B. cereus was found in 23.3% (47) of the food samples. The hemolytic enterotoxin hblA, hblC, and hblD were detected in 40.4%, 36.5%, and 75% of the isolates, respectively. Combination of all three hbl genes was detected in 19 isolates (36.5%). The nonhemolytic enterotoxin nheA, nheB, and nheC were detected in 61.5%, 92.3%, and 92.3% of the isolates, respectively. All three nhe genes were detected in 27 isolates (51.9%). The six enterotoxin genes (hemolysin BL [HBL] and NHE complexes) were detected in 25% of isolates. The cytK gene was detected in 53.8% of B. cereus isolates, whereas the bceT gene was detected in 69.2%. Only four emetic toxin-producing isolates were found to carry all the three genes of NHE complex, but none of the HBL complex ones. The B. cereus Enterotoxin-Reversed Passive Latex Agglutination assay results were strongly correlated with molecular detection of the L2 part of the HBL complex (p≤0.01). This study demonstrated the occurrence of potentially enterotoxic and emetic toxic B. cereus isolates in the food samples analyzed from Jordan.
Introduction
Toxin-producing B. cereus plays an important role as the causative agent of two different types of food poisoning: the emetic and the diarrheal types, which in most cases are relatively mild and self-limited (Ehling-Schulz et al., 2006). In some cases, it may be associated with a variety of nongastrointestinal infections such as bacteremia, devastating endophthlamitis, septicemia, meningitis, cellulites, abscess formation, endocarditis, osteomyelitis, upper and lower urinary tract infection, and pulmonary infection (Beecher and Wong, 1994). Additionally, B. cereus toxins can be of serious or even life-threatening infections especially in premature neonates, drug addicts, and immune-compromised patient (Drobniewski, 1993; Thaenthanee et al., 2005).
The emetic syndrome is caused by a single heat-stable toxin identified as cereulide produced by vegetative cells growing in the food (Granum, 2001), and leading to nausea and vomiting that develop immediately after the ingestion of food containing the toxin (Agata et al., 2002). The diarrheal syndrome is caused by a heat-labile enterotoxin produced during the vegetative growth of the bacteria in the small intestine, resulting in abdominal pain and watery diarrhea (Agata et al., 2002; Ehling-Schulz et al., 2004). Three enterotoxins are known to be associated with the diarrheal syndrome: the hemolytic enterotoxin hemolysin BL (HBL), nonhemolytic enterotoxin (NHE), and cytotoxin K (CytK) (Al-Khatib et al., 2007).
Recently, polymerase chain reaction (PCR) has been increasingly used for the detection of B. cereus genes encoding enterotoxins and emetic toxin and has proved to be a fast and sensitive method for the detection of these bacterial toxins (Al-Khatib et al., 2007).
In this study, we used PCR to characterize B. cereus isolates from food in Jordan including enterotoxin HBL and NHE complexes and CytK.
Materials and Methods
Collection of food samples
A total of 202 food samples were randomly collected from August 2008 to January 2009 from different local restaurants and supermarkets in different regions in Jordan (Amman, Irbid, and Al-Zarqa). The samples included cooked rice, yoghurt, spices, cake, pasta, raw meat, mashed potato, dried soup, and mixed salad. Samples were collected in the form as provided to the consumers, transported in an ice box directly to the laboratory, and examined within 2 h from the time of collection (Harmon et al., 1992). Additionally, extra five B. cereus isolates recovered from powder milk samples provided by the Jordan Food and Drug Administration/Amman were included in the study.
Isolation of B. cereus from food samples
To detect any B. cereus cells (vegetative or spores) present in the samples, 10 g of each food sample was homogenized manually in 90 mL of sterile 0.1% peptone water (1:10 dilution) and mixed well. One milliliter of food homogenate was transferred to 9 mL trypticase soy polymixin broth, and was incubated at 37°C for 48 h. One hundred microliters of the enriched sample was surface-plated on pre-dried polymyxin-egg yolk-mannitol-bromothymol-agar plates (Oxoid), and were incubated aerobically at 37°C for 24 h (Harmon et al., 1992). Two to three colonies suspected to be B. cereus (blue colonies surrounded by a zone of egg yolk precipitate) were picked up from each plate, streaked on blood agar plates, and incubated aerobically at 37°C for 24 h. Isolates forming β-hemolytic colonies and Gram-positive, rod-shaped bacteria with terminal to sub terminal spores without swelled sporangium were identified as Bacillus spp. of morphologic group 1. Isolates were confirmed as B. cereus based on Voges-Proskauer test, nitrate reduction test, citrate utilization, urea hydrolysis, indol production, motility, hemolytic activity, anaerobic growth, and the production of proteinaceous toxin crystals. B. cereus strain (ATCC 14579) was used throughout the study as a positive control (Shinagawa, 1990).
Detection of the HBL L2 component
Production of the L2 component of HBL by B. cereus was tested in culture filtrates using the B. cereus Enterotoxin-Reversed Passive Latex Agglutination (BCET-RPLA; Oxoid) kit as described by the manufacturer. Validity of the kits was confirmed by using positive and negative control.
Isolation of DNA
Genomic DNA for PCR was purified from bacterial cells cultured in brain heart infusion broth using commercial kit, and following the protocol provided by the manufacturer (Promega). The concentration of the extracted DNA was measured using SmartSpect™3000 (Bio-Rad). DNA samples were stored at −20°C until used.
Detection of enterotoxin genes and emetic toxin gene using PCR
The primers used for the detection of all studied genes with the anticipated size of the amplified product are presented in Table 1. For all assays, PCR mixture contained 12.5 μL of PCR master mix (Promega), 1 μmol of each primer, and 5 pg of DNA in a total volume of 25 μL of nuclease-free water. PCR was carried out on Brinkmann Gradient Eppendorf Mastercycler. Detection of HBL complex, NHE complex, and enterotoxin T genes was performed as reported by (Hansen and Hendriksen, 2001) with slight modification. The cycling conditions were denaturation at 94°C for 2 min, followed by 35 cycles of denaturation at 94°C for 15 sec, annealing at 55°C for 45 sec, extension at 72°C for 2 min, and a final extension at 72°C for 5 min. Detection of the cytK gene was performed as reported by Ngamwongsatit et al. (2008). The cycling conditions were denaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 45 sec, annealing at 54°C for 1 min, extension at 72°C for 2 min, and a final extension at 72°C for 5 min. Amplification using DNA from B. cereus ATCC 14579 was used as a positive control, whereas amplification using DNA of E.coli ATCC 25922 was used as a negative control. Detection of emetic toxin gene was performed as described by Ehling-Schulz et al. (2004). The cycling condition were denaturation at 95°C for 15 min, followed by 40 cycles of denaturation at 95°C for 30 sec, annealing at 60°C for 30 sec, extension at 72°C for 1 min, and a final extension at 72°C for 5 min. For all genes, the PCR products were detected in 1.5% agarose gel stained with ethidium bromide.
Statistical analysis
Data were analyzed using the computerized statistical program Statistical Package for Social Science (SPSS, version 15.0). p-Values less than 0.05 were considered significant.
Results
Prevalence of B. cereus in food samples
The qualitative distribution of the B. cereus isolates is shown in Table 2. Out of the 202 food samples analyzed, 47 samples (23.3%) contained B. cereus. High prevalence of B. cereus was detected in cake (50%), mixed salad (34.5%), pasta (33.3%), and yoghurt (31.8%) samples, whereas lower prevalence of B. cereus isolates was detected in cooked rice (20%), dried soup (20%), raw meat (16.7%), and spices (6.7%).
B. cereus toxin-encoding genes detection in food isolates
Prevalence of enterotoxins and emetic toxins genes of B. cereus isolated from food items is shown in Figure 1. The presence of the hblA gene encoding for the B part of HBL was detected in 21 isolates (40.4%), for the L1 part of HBL (hblD) in 39 isolates (75%), and for the L2 part (hblC) in 19 isolates (36.5%), respectively. Combination of the three hbl genes was detected in 19 isolates (36.5%), whereas the nheA gene encoding for the A part of the NHE complex was detected in 32 isolates (61.5%), the nheB gene encoding for the B part in 48 isolates (92.3%), and the gene for the C part of NHE complex (nheC) in 48 isolates (92.3%), respectively. All three nhe genes were detected in 27 isolates (51.9%). The cytK gene encoding for CytK was detected in 28 isolates (53.8%), and the bceT gene encoding for enterotoxin T was detected in 36 isolates (69.2%). The prevalence of emetic toxin-producing B. cereus isolates was 7.7% of the isolates.
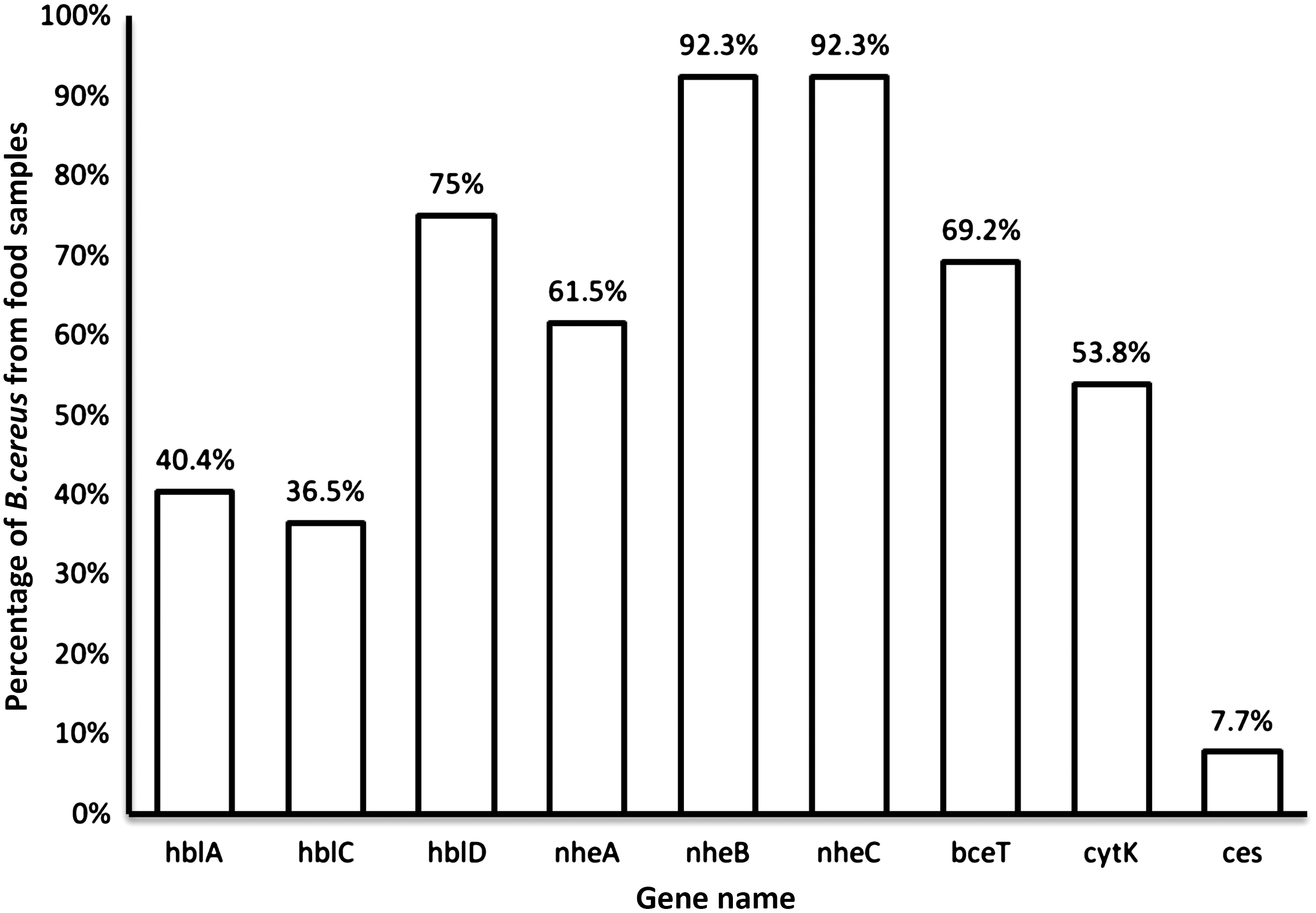
The prevalence of enterotoxin and emetic toxin genes in Bacillus cereus isolated from food samples in Jordan.
Toxin gene profiles of B. cereus isolated from various food items
B. cereus isolates were divided into five groups, according to the presence of hblCDA, nheABC, and cytK genes (Table 3). The hblCDA and nheABC were detected in 25% of isolates, among which 38% lacked cytK. The hblCDA or nheABC and/or cytK genes were detected in 40.4% of isolates. Isolates that carried some of the hblCDA and nheABC constituted 34.6% of the total, of which 33.5% carried cytK gene.
Miscellaneous food samples contain two isolates from spices, two isolates from pasta, one isolate from raw meat, and one isolate from dried soup.
The prevalence of the individual enterotoxin and emetic toxin genes in B. cereus isolated from various food items is shown in Table 4. High prevalence of both nheB and nheC (92%) was detected for each, whereas low prevalence was observed for hblA and hblC (40.4% and 36.5%, respectively). However, the cytK gene was detected in approximately half of the isolates. The bceT gene was detected in 69.2%, whereas the ces− gene was detected only in 7.7% of isolates (Table 4).
Miscellaneous food samples contained two isolates from spices, two isolates from pasta, one isolate from raw meat, and one isolate from dried soup.
Detection of the HBL L2 component
The production of the L2 component of HBL complex by the 52 B. cereus isolates was analyzed using the Oxoid RPLA test. A total of 18/19 (94.7%) of the B. cereus isolates were positive in both BCET-RPLA assay and PCR test. The L2 titers ranged between 64 and ≥128 ng/mL; however, 52.6% of the isolates had a titer ≥128 ng/mL. Out of the 19 isolates, 1 was positive for the hblC gene but negative by BCET-RPLA assay. Five isolates that were negative for hblC gene by PCR reacted positively in the RPLA assay with a titer range between 8 and 32 ng/mL. These five isolates were found to carry both hblA and hblD genes, and only the hblD gene. Significant correlation between the BCET-RPLA assay and PCR test for the detection of the L2 part of the HBL complex was observed (p≤0.01) using the Fisher's Exact Test.
Discussion
The results of this study showed a relatively high prevalence (23.3%) of B. cereus in the food samples examined. The highest prevalence of B. cereus was observed in cake samples (50%) whose formulation mainly includes wheat flour and cream. Both of them can be considered as a source of contamination by B. cereus (Wong et al., 1988; Iurlina et al., 2006). The prevalence of B. cereus in mixed salad and pasta samples collected from local restaurants was 34.5% and 33.3%, respectively. Mixed salad and pasta consisted mainly of different types of vegetables such as maize, carrot, potato, pepper, and lettuce. B. cereus, as spore-forming bacteria, are normally present in soil and water, and easily spread to food, especially those of plant origin (Granum, 2001). High prevalence of B. cereus was detected in milk powder and yoghurt samples (31.8%), which could be attributed to the fact that pasteurization is insufficient to kill the spores that are very hydrophobic and can adhere to the surfaces of the pipelines of the dairy industry (Anderson et al., 1995, 1998). Further, yogurt and other dairy products are shop-made from fresh sheep, goat, and cow milk on small scale that may be contaminated with B. cereus spores. In accordance to previous studies (Granum et al., 1996; Iurlina et al., 2006), B. cereus was observed in spices, dried soup, and cooked rice (6.7%, 20%, and 20%, respectively). Contamination of cooked rice may be due to the presence of B. cereus spores in uncooked rice and spices used in the preparation of these products (Sarrías et al., 2002). The percentages of contamination of the different food products observed in the current study are within the range of what were observed in similar studies, where Iurlina et al. (2006) reported that the prevalence of B. cereus in food products like honey, flour, cheese, and spices was from 23% to 50%. Similarly, Reyes et al. (2007) reported that the prevalence of B. cereus ranged between 34.5% and 62.5% among various dried milk products such as milk powder, pudding milk, and milk with rice.
B. cereus has been recognized as a causative agent of food poisoning with diarrheal or emetic syndromes, and a variety of nongastrointestinal infections such as bacteremia, meningitis, and post-traumatic eye infections (Thaenthanee et al., 2005). Different toxins are responsible for the gastrointestinal diseases caused by B. cereus. At least three enterotoxins are associated with the diarrheal syndrome (HBL, NHE, and CytK), whereas the emetic syndrome is caused by a single toxin called cereulide (Ehling-Schulz et al., 2006). In this study hblA, hblC, and nheA genes were detected in <50% of B. cereus isolates (range 36.5%–40.5%). This value is lower than those previously reported from other countries (50%–86%) using similar methods (Mäntynen and Lindstrom, 1998; Hansen and Hendriksen, 2001; Al-Khatib et al., 2007). The hblD, nheB, and nheC genes were detected in 75%, 92.3%, and 92.3%, respectively. Our results are in agreement with the results of other studies (60%–95%) from food and foodborne outbreaks (Hansen and Granum et al., 1996; Hendriksen, 2001; Guinebretière et al., 2002; Al-Khatib et al., 2007). Similarly, the prevalence of the cytK gene (53.8%) and the bceT gene (69.2%) was within the range observed previously (Granum et al., 1996; Hsieh et al., 1999; Ngamwongsatit et al., 2008). Our results showed that 36.5% of the B. cereus isolates carried the three hbl genes (hblA, hblC, and hblD). Further, 51.9% of the B. cereus isolates contained the three nhe genes (nheA, nheB, and nheC), and 25% of the B. cereus isolates carried the six enterotoxin genes (HBL and NHE complexes). The observed genetic heterogeneity among B. cereus strains might be associated with sequence polymorphism more than the lack of one of the genes composing the hbl or nhe operon (Guinebretière et al., 2002).
All B. cereus isolates in this study possessed hemolytic activity on blood agar plates, although not all of them were PCR positive for the hemolytic enterotoxin (HBL complex). These results are in agreement with what was reported for by Thaenthanee et al. (2005). The hemolytic activity of B. cereus might be toxins/virulence factors other than that encoded by HBL genes (Thaenthanee et al., 2005). Thus, the results of current study provide more support for this hypothesis.
In this study, the five isolates that were negative for the hblC gene by PCR showed positive reactions in the RPLA assay. These five isolates carried both hblA and hblD genes, or carried only the hblD one. On the other hand, 1 of the 19 isolates was positive for the hblC gene but negative for BCET-RPLA assay. These findings could be explained on the fact that the RPLA test is not specific for the L2 part only, but the gene sequence variations that might also affect protein antigenicity can lead to false-negative results in both PCR test and BCET-RPLA assay (In't Veld et al., 2001; Thaenthanee et al., 2005).
In conclusion, this study suggests the occurrence of potentially enterotoxic and emetic toxic B. cereus in the food samples analyzed from Jordan, with some variations in genotypes prevalence among the isolates.
Footnotes
Acknowledgments
This work was performed with funds from the Faculty of Scientific Research in Jordan University of Science and Technology Grant Number 137/2008.
Disclosure Statement
No competing financial interests exist.