
Abstract
A protective digestive microflora helps prevent and reduce broiler infection and colonization by enteropathogens. In the current experiment, broilers fed diets supplemented with probiotics and essential oil (EO) blends were infected with a standard mixed Eimeria spp. to determine effects of performance enhancers on ileal and cecal microbial communities (MCs). Eight treatment groups included four controls (uninfected-unmedicated [UU], unmedicated-infected, the antibiotic BMD plus the ionophore Coban as positive control, and the ionophore as negative control), and four treatments (probiotics BC-30 and Calsporin; and EO, Crina Poultry Plus, and Crina PoultryAF). Day-old broilers were raised to 14 days in floor pens on used litter and then were moved to Petersime batteries and inoculated at 15 days with mixed Eimeria spp. Ileal and cecal samples were collected at 14 days and 7 days postinfection. Digesta DNA was subjected to pyrosequencing for sequencing of individual cecal bacteria and denaturing gradient gel electrophoresis (DGGE) for determination of changes in ileal and cecal MC according to percentage similarity coefficient (%SC). Pyrosequencing is very sensitive detecting shifts in individual bacterial sequences, whereas DGGE is able to detect gross shifts in entire MC. These combined techniques offer versatility toward identifying feed additive and mild Eimeria infection modulation of broiler MC. Pyrosequencing detected 147 bacterial species sequences. Additionally, pyrosequencing revealed the presence of relatively low levels of the potential human enteropathogens Campylobacter sp. and four Shigella spp. as well as the potential poultry pathogen Clostridiun perfringens. Pre- and postinfection changes in ileal (56%SC) and cecal (78.5%SC) DGGE profiles resulted from the coccidia infection and with increased broiler age. Probiotics and EO changed MC from those seen in UU ilea and ceca. Results potentially reflect the performance enhancement above expectations in comparison to broilers not given the probiotics or the specific EO blends as feed supplements.
Introduction
Supplementing young chicks with beneficial bacteria enhance early establishment of a protective MC and live performance (Nurmi and Rantala, 1973; Lloyd et al. 1977; Barnes et al., 1980). The growing recognition of the importance of healthy MC has resulted in increased interests in bacterial probiotic dietary supplements (Blankenship et al., 1993; Nisbet et al., 1993; Schneitz et al., 1998; Flint and Garner, 2009). Probiotic supplements offer early protection against enteropathogen invasion and colonization, can stimulate immune response, and may take advantage of nutritive components unavailable to the host.
In the current report, the effects of two commercial probiotics and two commercial specific EO on ileal and cecal MC in broiler chickens were evaluated before and after a mixed Eimeria spp. infection. Two molecular techniques, pyrosequencing and denaturing gradient gel electrophoresis (DGGE), were applied to monitor the impact of treatments and Eimeria infection on MC. The combined analyses of the two techniques were applied to give views at two levels of community changes. Pyrosequencing was applied to examine the impact of treatment on pre- and postinfection cecal on individual bacteria. DGGE was used for more gross comparisons of treatments on pre- and postinfected ileal and cecal MC.
Materials and Methods
Broiler husbandry
Day-old male Ross 708 chickens (384) were randomly placed in 48 floor pens (8/pen), each with litter used by broilers given the same treatments in two previous experiments. Litter management was applied to simulate commercial production systems. Downtime between experiments was 12 days. No additional bedding material was added. Pens were distributed to have an empty pen without chickens and without litter between pens to avoid cross contamination. Litter management between flocks was done with the same tools in all pens of similar treatment, and equipment was cleaned and disinfected before handling litter in pens assigned to different treatments. Daily biosecurity measurements were taken to avoid cross contamination among pens or cages. From 1 to 14 days, personnel did not enter the pens. Personnel did not have to enter pens for routine management, and disposable boots and gloves were used for pens, equipment, birds, feed, and samples assigned different treatments.
Chickens were distributed in eight treatments with six replicates each. Broilers were given balanced feed (NRC, 1994) and water provided ad libitum. At 14 days of age, 288 broilers (36/treatment) were transferred to Petersime batteries (6/cage). Lighting simulated commercial conditions: 1 to 8 days, 23 h of light/day at 2.0 foot-candles, 1 h of darkness between 2 and 3 am; and 8 to 22 days, 6 h of darkness between 9:00 pm and 3:00 am. Rearing was according to safe practices authorized by the Animal Use Protocol Committee.
Diets and treatments
The eight treatments (Table 1) included four controls and four treatment diets. Broilers were fed starter crumbles to 22 days of age. All diets contained the phytase enzyme Ronozyme P CT (DSM Nutritional Products, Basal, Switzerland) at 168 g/ton. Diets also contained vermiculite as filler to allow replacement by either phytase, or enough dicalcium phosphate to add 0.1% phosphorus in treatment Crina Poultry AF 100 ppm (CPF) (Table 1), or probiotics or the specific EO blend according to treatment distribution.
AGP, antibiotic growth promoter.
Coccidia
All groups except uninfected-unmedicated control (UU) were given at 15 days of age a standard oral mixed Eimeria inoculum of sporulated oocysts: E. acervulina (2×105/mL), E. maxima (2.5×104/mL), and E. tenella (5×104/mL) (provided by Dr. Mark C. Jenkins, Animal Parasitic Diseases Laboratory, Agricultural Research Service, U.S. Department of Agriculture). Oocytes were susceptible to ionophore.
Sample collection and DNA extraction
Ileal and cecal digesta were collected from six broilers per group pre-Eimeria infection at 14 days of age and from six broilers per group postinfection at 22 days of age. Samples were lyophilized and stored at −80°C until DNA extraction. Lyophilized samples (0.25 g) were suspended in 180 μL of lysis buffer (QIAmp DNA Mini Kit; QIAgen Incorporated, Valencia, CA) containing 20 mg/mL of lysozyme and extracted according to kit instructions.
Pyrosequencing massively parallel bacterial tag-encoded FLX amplicon pyrosequencing
Bacterial tag-encoded FLX amplicon pyrosequencing was performed as described previously using Gray28F 5′TTTGATCNTGGCTCAG and Gray519r 5′GTNTTACNGCGGCKGCTG (Bailey et al., 2010a,b; Callaway et al., 2010; Finegold et al., 2010; Gontcharova et al., 2010a,b; Olafson et al., 2010; Pitta et al., 2010; Smith et al., 2010; Stephenson et al., 2010; Williams et al., 2010; Andreotti et al., 2011; Handl et al., 2011; Ishak et al., 2011) with primers numbered relative to Escherichia coli 16S rDNA. Initial generation of the sequencing library utilized a one-step polymerase chain reaction (PCR) with a total of 30 cycles, a mixture of HotStart and HotStar high fidelity taq polymerases (Qiagen), and amplicons originating and sequencing extending from the 28F with average read length of 400 bp. Tag-encoded FLX amplicon pyrosequencing analyses utilized Roche 454 FLX instrument with Titanium reagents, titanium procedures performed at the Research and Testing Laboratory (Lubbock, TX) based upon RTL protocols (
Pyrosequence bacterial diversity data analysis
After sequencing, all failed sequence reads, low quality sequence ends and tags, and primers were removed and sequence collections depleted of any nonbacterial ribosome sequences and chimeras using B2C2 (Gontcharova et al., 2010b) as previously described (Bailey et al., 2010a,b; Finegold et al., 2010; Gontcharova et al., 2010a; Olafson et al., 2010; Pitta et al., 2010; Smith et al., 2010; Stephenson et al., 2010; Williams et al., 2010; Andreotti et al., 2011; Handl et al., 2011; Ishak et al., 2011). To identity the remaining bacterial sequences, sequences were denoised, assembled into clusters, and queried using a distributed BLASTn .NET algorithm (Dowd et al., 2005) against a high-quality 16S bacterial sequence database (based upon similar criteria utilized by RDP ver 9 (Cole et al., 2009)) derived and curated monthly from NCBI (01-01-11). Using a .NET and C# analysis pipeline, the resulting BLASTn outputs were compiled, validated using taxonomic distance methods, and data reduction analysis performed (Bailey et al., 2010a,b; Callaway et al., 2010; Finegold et al., 2010; Gontcharova et al., 2010a,b; Olafson et al., 2010; Pitta et al., 2010; Smith et al., 2010; Stephenson et al., 2010; Williams et al., 2010; Andreotti et al., 2011; Handl et al., 2011; Ishak et al., 2011).
Pyrosequence bacterial identification
Based upon the above BLASTn-derived sequence identity (percent of total length query sequence that aligns with a given database sequence) and validated using taxonomic distance methods, the bacteria were classified at the appropriate taxonomic levels based upon the following criteria. Sequences with identity scores, to known or well-characterized 16S sequences, >97% identity (<3% divergence) were resolved at the species level, between 95% and 97% at the genus level, between 90% and 95% at the family, and between 85% and 90% at the order level, 80% and 85% at the class, and 77% to 80% at phyla. Evaluations presented at each taxonomic level, including percentage compilations, represent all sequences resolved to their primary identification or their closest relative (Bailey et al., 2010a,b; Callaway et al., 2010; Finegold et al., 2010; Gontcharova et al., 2010a,b; Olafson et al., 2010; Pitta et al., 2010; Smith et al., 2010; Stephenson et al., 2010; Williams et al., 2010; Andreotti et al., 2011; Handl et al., 2011; Ishak et al., 2011).
Denaturing gradient gel electrophoresis
Extracted DNA adjusted to 42 ng/μL and 1 μL from each of six samples per treatment were pooled for PCR (Zhou et al., 2007), targeting the 16S rDNA gene V3 region. Reactions were performed in 25 μL according to Muyzer et al. (1993) with modification (Hume et al., 2003). Electrophoresis was conducted using a DCode Universal Mutation Detection System (Bio-Rad Laboratories, Hercules, CA). Band pattern relatedness was determined with Molecular Analysis Fingerprinting Software, version 1.6 (Bio-Rad Laboratories), based on the Dice similarity coefficient (SC) and unweighted pair group method using arithmetic averages (UPGMA) for clustering analysis and comparison according to percentage SC (%SC).
Statistical analysis
The DGGE profiles were digitized and converted to a binary matrix using a script (available on request) written in the Python programming language (Python Software Foundation, Hampton, NH). Taxonomic assignment of pyrosequencing reads was carried out using BLAST. The annotated sequences were used to calculate the relative percent abundance of taxa per sample. The pyrosequencing abundance matrix and the DGGE binary matrix were further analyzed with the R statistical software (Foundation for Statistical Computing, Vienna, Austria;
Results
Pyrosequencing
Nine bacteria phyla were detected among the 48,461 sequences identified in cecal samples taken from eight treatment groups in Eimeria pre- and postinfection broilers: Actinobacteria, Bacteroidetes, Candidatus Poribacteria, Cyanobacteria, Firmicutes, Fusobacteria, Proteobacteria, Tenericutes, and Verrucomicrobia. Bacteroidetes, Firmicutes, and Proteobacteria are represented by at least 1% of identified sequences (Tables 2 and 3). Orders Bacteroidales, Clostridiales, Bacteriales, and Enterobacteriales were detected in preinfection ceca, whereas postinfection orders were Clostridiales, Bacteroidales, Campylobacterales, Lactobacillales, and Enterobacteriales. Further classification of sequences identified, including those at <1%, revealed 15 classes, 20 orders, 44 families, 90 genera, and 147 species.
UU, unmedicated-uninfected control; UI, unmedicated-infected control; PC, positive control; NC, negative control; BC30, BC-30 probiotic; CAL, calsporin probiotic; CPP, Crina Poultry Plus 300 ppm; CPF, Crina Poultry AF 100 ppm; nd, not detected.
Seven species representing at least 1% of sequences were detected at preinfection (Table 2), whereas 12 were detected at postinfection (Table 3). Some sequences are listed as genus unnamed species to designate these as potentially novel sequences unmatched in the data base. No specific pattern of predominance was seen across treatments. However, Clostridium sp., Papillibacter sp., Ruminococcus sp., and Lactobacillus sp. were detected at relatively high numbers. There was no pattern for species relative abundance in Eimeria postinfection ceca (Table 3). Every postinfection group had at least one sequence hit at or above 10%.
Potential human enteropathogens were detected at pre- and postinfection. A Campylobacter sp. was detected in all groups except UU at postinfection. Shigella boydii, S. dysenteriae, S. flexneri, S. sonni, and an unidentified Shigella sp. were detected. The Shigella boydii and S. flexneri were not detected at preinfection. The poultry pathogen Clostridiun perfringens was detected at postinfection in unmedicated-infected control (UI), BC30, and Crina Poultry Plus 300 ppm (CPP) (data not shown).
PCA (Fig. 1) of pyrosequence data and body weight (BW) shows the relationship among cecal MC at pre- and postinfection. Distances between samples correlate with increasing MC dissimilarity. The first axis explained 31.4% and second axis 21.9% of variation. The first axis corresponded to Eimeria infection (p=0.001), revealing two groups: preinfection (circles) and postinfection (squares). Significant correlation was observed between MC composition and the relative percent average BW (p=0.0001, R 2=0.9).
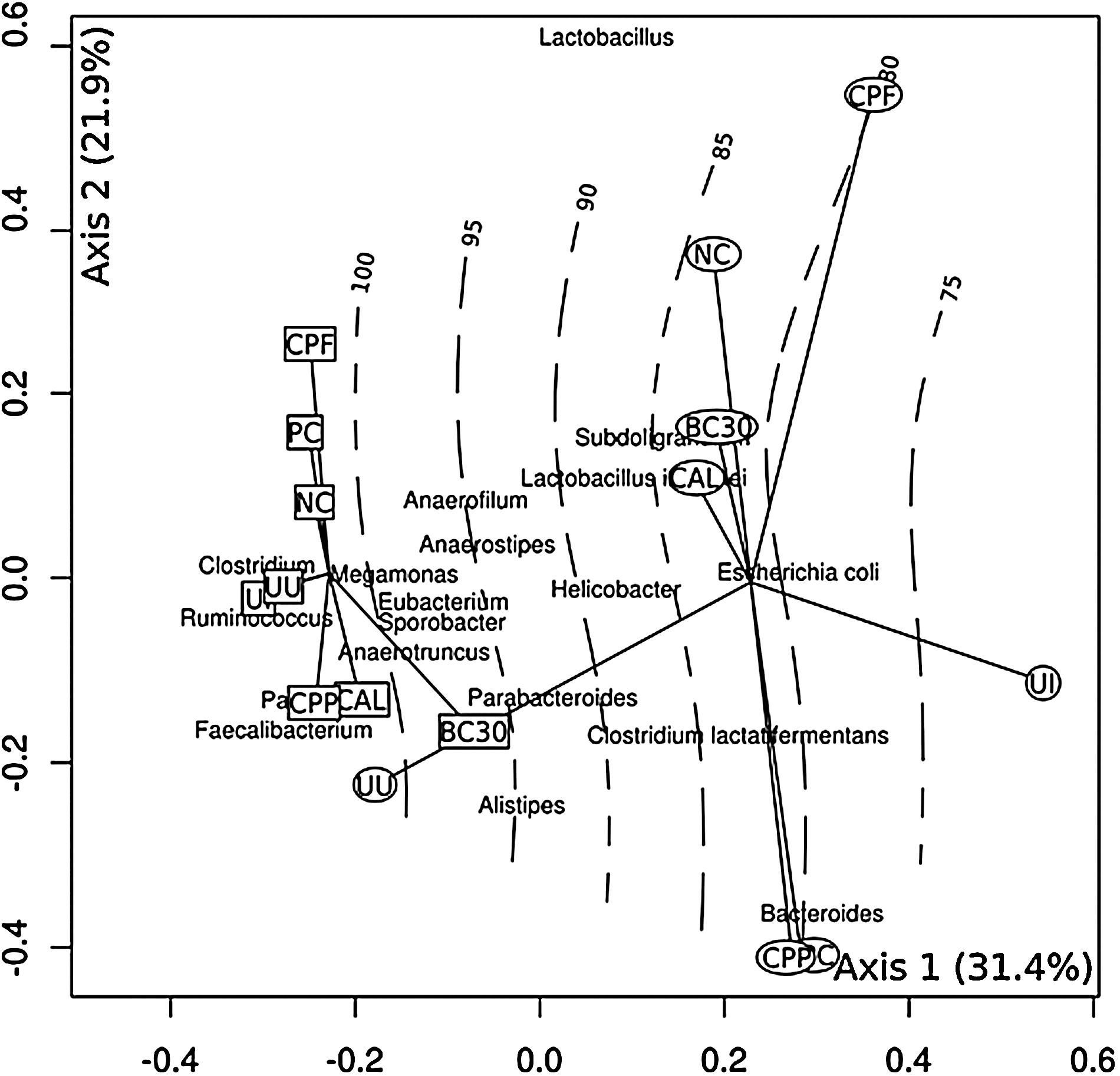
Principal component analysis ordination plot of cecal bacterial microflora identified in pre- (squares) and post-Eimeria (circles) infection ceca. Solid lines connect samples into pre- and postinfection groups (centroids). Axes 1 and 2 explain 53.3% of the variation present in the data. Contour lines represent the relative percent average body weight data fit to samples ordination scores. Taxa names indicate the most abundant taxa occuring at different locations in ordination space. The difference between the groups (centroids) is significant (p<0.001). Abbreviations inside the symbols represent treatments as indicated in Table 1.
Denaturing gradient gel electrophoresis
The ileal DGGE MC dendrogram shows two main groups composed mainly of pre- and postinfection MC, respectively (Fig. 2). However, postinfection UU clustered with the preinfection MC and exhibiting 78.9%SC with negative control (NC) and Calsporin. The NC and Calsporin MC shared 87.9%SC, whereas UI clustered with postinfection groups. The preinfection UI exhibited very close alignment with the postinfection NC, positive control (PC), UI, and BC30.

Dendrogram of denaturing gradient gel electrophoresis comparing 16S rDNA amplicon patterns of ileal (Il) contents bacterial populations in chicks at 14 days of age pre-infection and at 22 days of age and 7 days after mixed Eimeria spp. postinfection. Relative similarity of band patterns is indicated by their grouping on the dendogram and the percentage similarity coefficient (bar). Abbreviations represent treatments as indicated in Table 1.
Cecal MC also segregated into mainly pre- and postinfection groups (Fig. 3). Postinfection UU clustered with preinfection MC and in close association with preinfection UI (91.8%) and UI (88.2%). The preinfection CPF clustered with the postinfection CPP (83.6%), whereas the postinfection CPF and UI exhibited MC most unlike the other groups (51.6% and 54.6%, respectively).
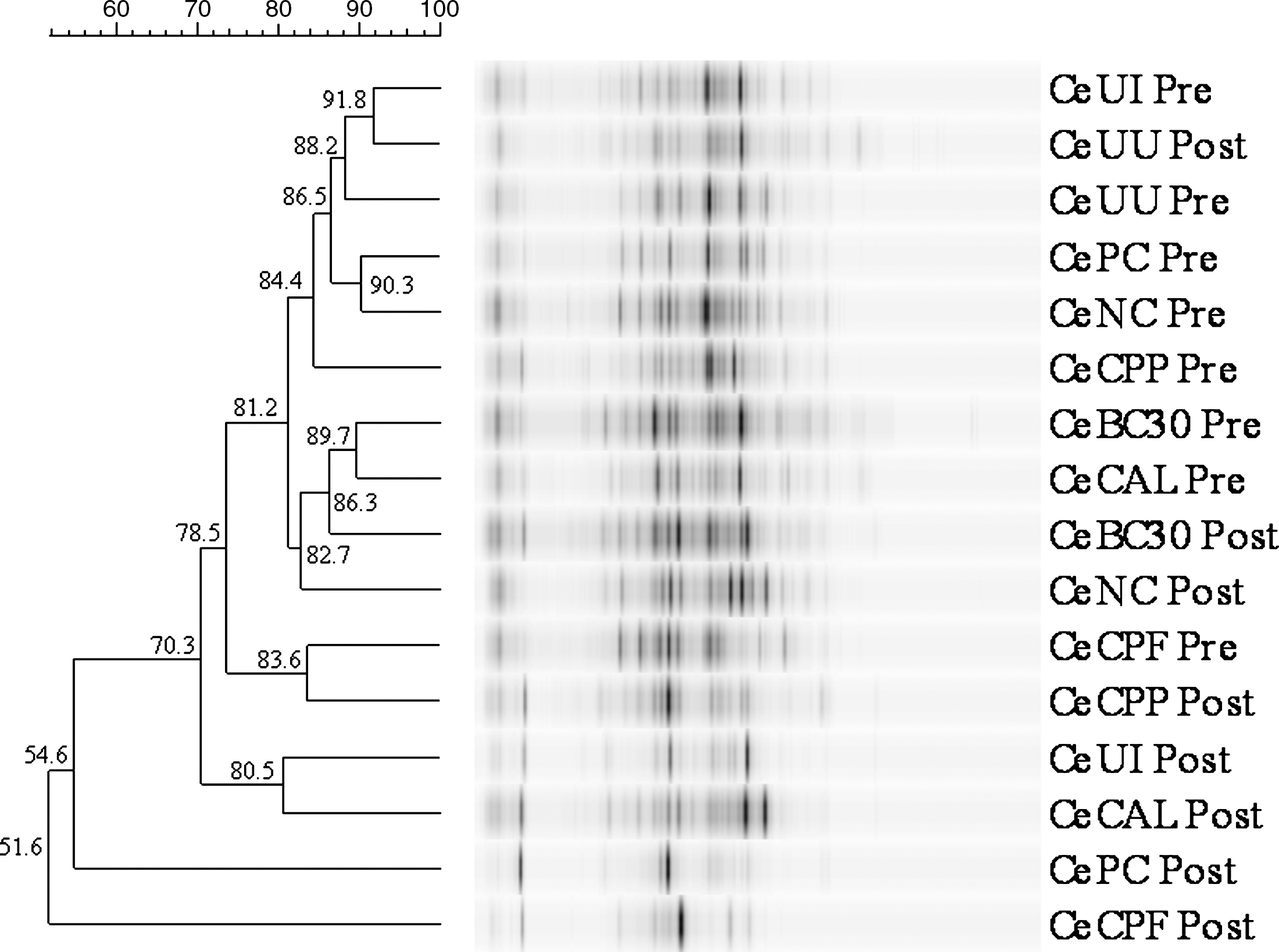
Dendrogram of denaturing gradient gel electrophoresis comparing 16S rDNA amplicon patterns of cecal (Ce) contents bacterial populations in chicks at 14 days of age pre-infection and at 22 days of age and 7 days after mixed Eimeria spp. postinfection. Relative similarity of band patterns is indicated by their grouping on the dendogram and the percentage similarity coefficient (bar). Abbreviations represent treatments as indicated in Table 1.
PCA revealed that Eimeria infection significantly (p<0.009) affected ileal MC (Fig. 4). Similar results were seen in the PCA of cecal MC (Fig. 5). The arrangement of groups shows that cecal MC at preinfection have significantly (p<0.04) different MC compared to postinfection broilers.
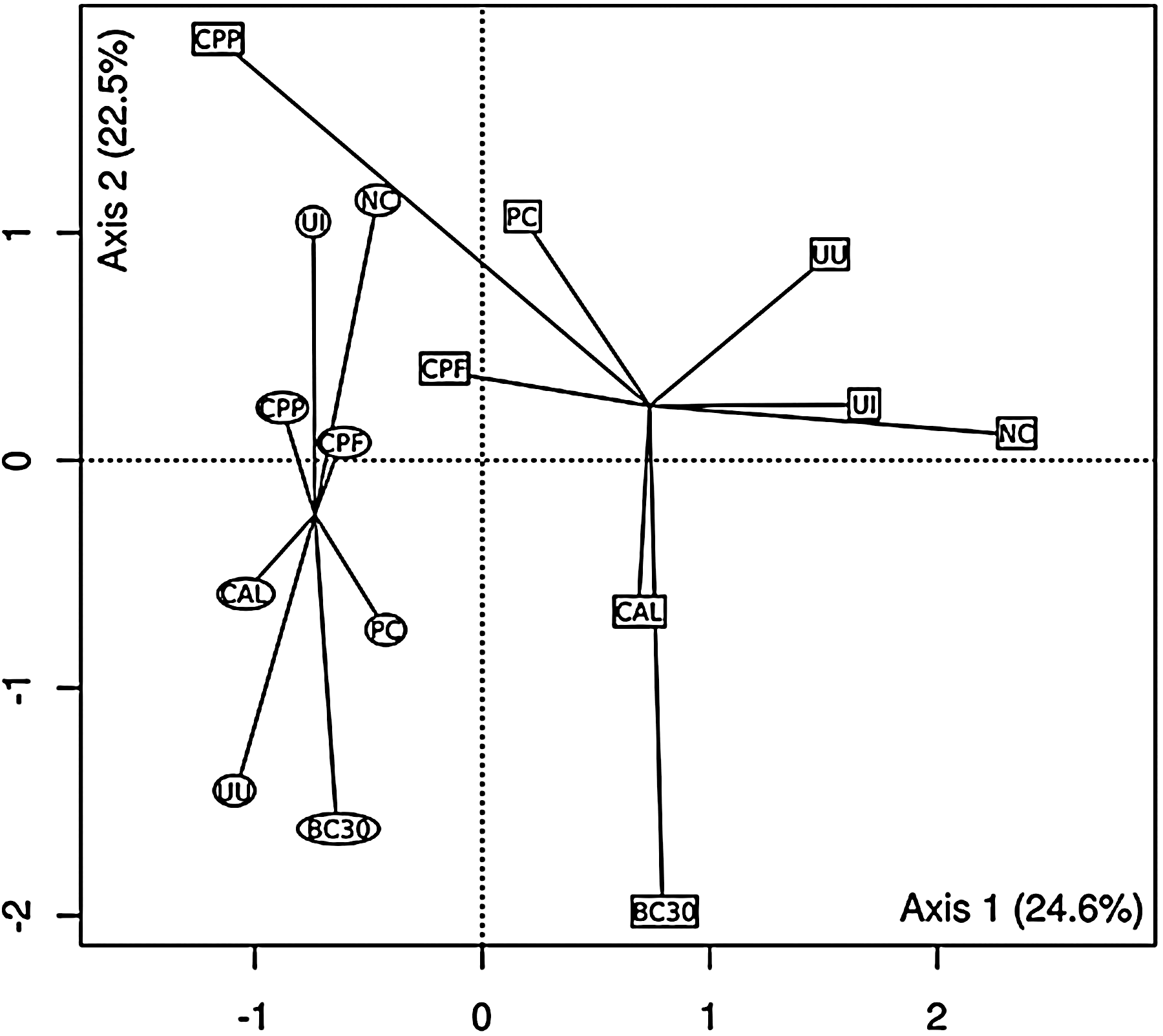
Principal component analysis ordination plot of denaturing gradient gel electrophoresis profiles from pre- (squares) and post-Eimeria (circles) infection ceca. Solid lines connect samples into pre- and postinfection groups (centroids). Axes 1 and 2 explain 47.1% of the variation present in the data. The difference in centroids is significant (p<0.009). A inside the symbols represent treatments as indicated in Table 1.
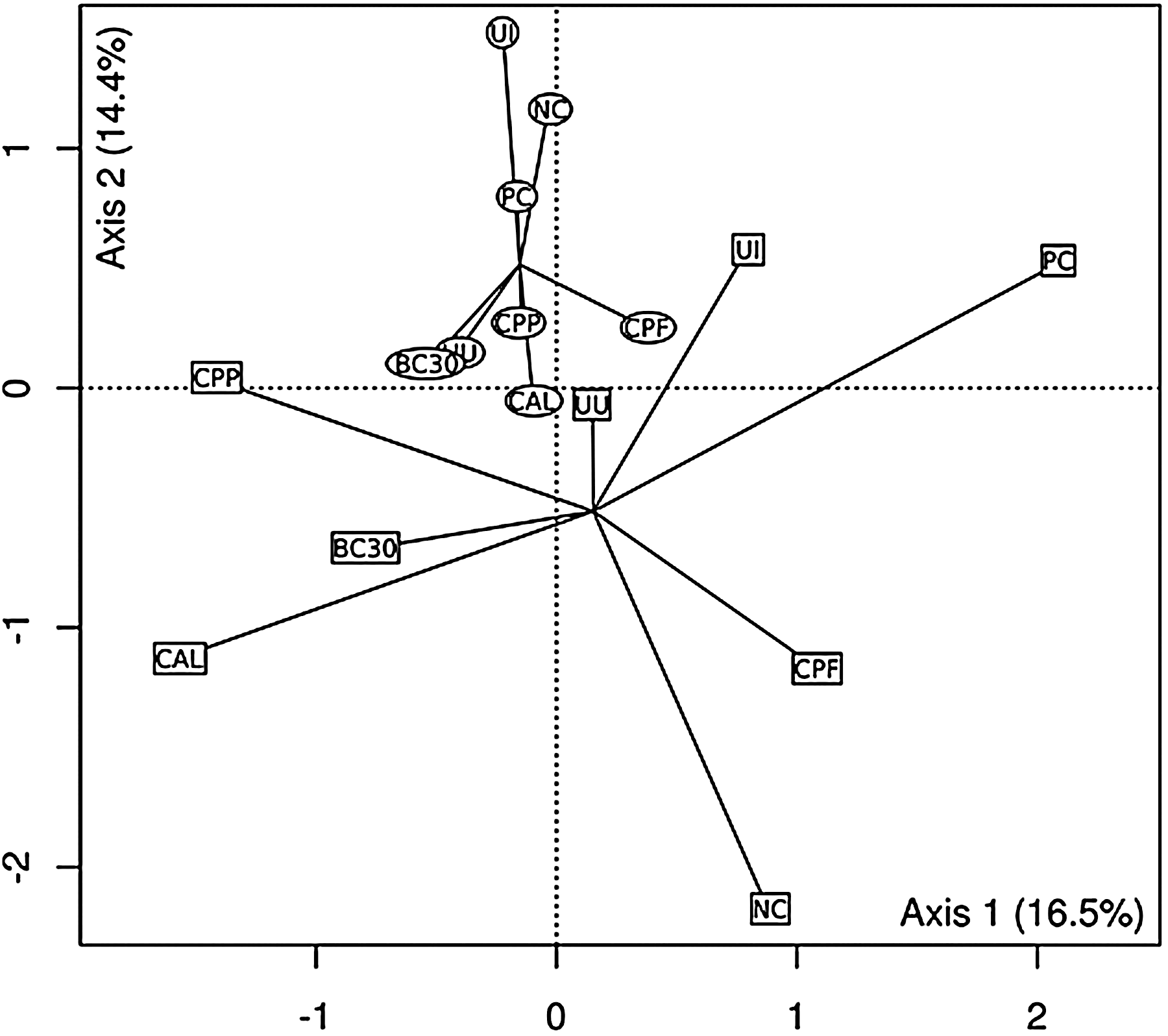
Principal component analysis ordination plot of denaturing gradient gel electrophoresis profiles from pre- (squares) and post-Eimeria (circles) infection ilea. Solid lines connect samples into pre- and postinfection groups (centroids). Axes 1 and 2 explain 30.9% of the variation present in the data. The difference in centroids is significant (p<0.04). Abbreviations inside the symbols represent treatments as indicated in Table 1.
Growth and feed performance
No real differences in treatments for body weight gain (BWG) or feed conversion ratio (FCR) were seen in broilers at 14 days of age (Table 4). By 22 days, group UU were significantly (p<0.05) more BWG than all other treatment groups, with the lowest BWG in UI, broilers given no treatment other than infection. The UU group also had better (p<0.05) FCR than UI, NC, and CPP and with a ratio numerically lower than all other groups. Broilers in UI, accordingly, had the worst FCR of all treatment groups.
All probiotic and essential oil treatments contained the same ionophore as PC and NC.
Means in rows with no common superscript differ significantly (p<0.05) by Tukey's test.
PC, positive control (antibiotic+ionophore); NC, negative control (ionophore only); CPP, Crina Poultry Plus, essential oil at 300 ppm; CPF, Crina Poultry AF, essential oil at 100 ppm; BWG, body weight gain; FCR, feed conversion ratio; CV, coefficient of variation.
Discussion
An objective of the current line of poultry MC studies is to identify bio-organisms and chemical agents that enhance production and off-set negative effects of pathogen colonization or stress conditions. Treatment effects in the present study seen as differences in DGGE banding patterns likely resulted from bactericidal, bacteriostatic, or beneficial effects of the probiotic Bacillus spp. or bioactive EO. The current study, to our knowledge, is one of the first to combine a broad-spectrum species molecular identification, with far less bacterial selection than traditional culturing, and the DGGE visual referencing. However, DGGE band identity is still dependent upon imprecise excision of closely positioned gel bands for PCR and database referencing. Alternatively, pyrosequencing per sample costs make DGGE more attractive as a treatment discriminating tool for MC studies.
Broiler digestive MC were influenced by two commercial probiotics and two EO. From pre- to postinfection, there was an increase in the number of species at 1% or more. This increase may be the result of broiler and MC maturation (Lu et al., 2003) as well as from treatments. However, DGGE demonstrated the impact of Eimeria infection on MC as seen with the clustering of postinfection UU with preinfection groups. Additionally, PCA contour lines, showing relative percent average BW, along with positioning of postinfection UU with preinfection groups, indicated the pronounced effect of infection on BW loss as well as shifts in MC.
Lactobacillus, Enterococcus, and Es. coli were detected at relatively high levels and are common commercial probiotic ingredients (Huys et al., 2006). In general, in vitro and in vivo oregano EO and BMD treatment reduce the numbers of these genera (Kivanc et al., 1991; Elgayyar et al., 2001; Burt and Reinders, 2003; Peñalver et al., 2005; Biggs and Parsons, 2008). Cecal Lactobacillus at preinfection was relatively high in CPP and CPF, whereas Lactobacillus was high in BC30 and CPF at postinfection. The lack of apparent inhibition of Lactobacillus at pre- and postinfection or Enterococcus in postinfection CPP may be due to differences in EO composition used in the current study as compared to others.
The commercial probiotics contained known Bacillus spp. However, an unidentified Bacillus sp. was detected below 1% in postinfection UU, BC30, and CPF and did not register as a Bacillus in the probiotics administered. This result is not surprising, since residence of introduced probiotic bacteria is often transient and at undetectable levels (Freter et al., 1983; Jin et al., 1998; Priyankarage et al., 2004).
In spite of the application of commercial probiotics and the presence of commonly applied probiotic bacteria, several potential human enteropathogens were detected, mainly at Eimeria postinfection. However, these enteropathogens were not detected or were at low numbers in UU broilers. Eimeria oocyte vaccination in 1-day-old broilers can render immune protection against Salmonella (Volkova et al., 2011). However, Eimeria infection in 2-week-old broilers can disrupt cecal MC (Hume et al., 2006; Oviedo-Rondón et al., 2006). This disruption appears sufficient to render broilers vulnerable to enteropathogen colonization. A low level Campylobacter colonization was seen at postinfection, but was not detected in UU and was above 10% in NC (given Coban). Usually, there is no association between coccidiostat treatment and increased susceptibility to enteropahtogen colonization.
While pyrosequencing is more useful at detecting minor changes in specific taxa, DGGE gives a qualitative profile of MC with sensitivity at 1% of the total community. Ileal and cecal DGGE profiles confirmed previous reports demonstrating the effects of Eimeria on MC (Hume et al., 2006; Oviedo-Rondón et al., 2006). Ileal MC separated into two main groups, pre- and postinfection. This partitioning is partially an expected result of broiler aging. Grouping of postinfected ileal UU with preinfected groups reflects that this group received no medication and no infection. Apparently, aging did not have as much effect on ileal MC as did treatments and Eimeria infection. However, preinfected UI was more similar to post- than to preinfected broilers. Visual examination of band patterns suggests that the main difference was not the band presence or absence, but, possibly, relative band intensity. Differences in intensities may reflect minor and natural variation in aging populations (Zhu et al., 2002; Hume et al., 2003). The greater similarity between postinfection than between preinfection groups suggests stabilization of MC with age and lessening of variation as MC mature. Of course, another consideration may be substantial influence exerted by Eimeria.
There were some similarities in cecal MC clustering as seen in ilea. Cecal postinfection UU profile was very similar to preinfection UU and UI. Grouping of cecal UU with UI is more likely than the same occurring in ilea due to the more stable, uniform cecal environment. Similar to what was seen in postinfection ilea, preinfection PC and NC, while grouped with UU and UI, reflect the lack of treatment with probiotic or specific EO on these populations.
Intestinal MC are vital to broiler performance and health. Eimeria infection effects on MC may be seen as changes in BWG and feed efficiency. At postinfection, BWG except in UU reflect the negative effects of infection. Poor performance is an expected result of Eimeria infection (Moynihan, 1950) and multiple approaches target conventional antimicrobials to reduce these negative effects. By day 22 in the current study, none of the feed additives reduced the negative effects of infection on BWG to levels seen in UU. However, all feed additives had some positive effect on decreased weight gain. Eimeria infection alters the MC in affected broilers (Hume et al., 2006; Oviedo-Rondón et al., 2006). The inference is that the debilitating effects of Eimeria infection were lessened by these fed additives. The ultimate goal is to identify specific effects of feed additives on beneficial and debilitating MC and to identify those MC components most supportive of broiler production.
Treatments including the two commercial probiotics and the two specific EO modified broiler ileal and cecal digestive MC. Pyrosequencing was very sensitive at detecting shifts in individual identified sequences, whereas DGGE was adept at detecting gross shifts in entire populations. These combined techniques offered added versatility toward unraveling the effects of Eimeria infection and performance enhancing feed additives on broiler MC. Results show modifications of MC resulting from treatments before infection with a standard mixed Eimeria spp. as well as the effects after mild infection. Pyrosequencing and DGGE offer individual advantages in addition to their respective sensitivity and ability to detect gross shifts in populations. The effects of infection and treatment with either performance enhancing or therapeutic antibiotics are well understood at the macro level. Efforts to uncover the contributions made by shifts in individual bacteria in the greater community, similarly as applied in the current study, will aid in the development of intervention strategies for protection and treatment against pathogen infection and colonization. Molecular techniques such as pyrosequencing, in contrast to the more bio-selective traditional culture techniques, offer a considerably more objective view of minor and major contributors to the general MC.
Disclaimer
Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Footnotes
Disclosure Statement
No competing financial interests exist.