
Abstract
Salmonella can live undetected in the gut of food animals and be transmitted to humans. Animal diets can impact intestinal populations of foodborne pathogens, including Salmonella spp. Orange juice production results in a waste product, orange peel and orange pulp, which has a high nutritive value and is often included in cattle diets as a least-cost ration ingredient. Here we show that the inclusion of orange peel products reduced Salmonella Typhimurium populations in the gut of experimentally inoculated sheep. Sheep (n=24) were fed a cracked corn grain-based high grain diet that was supplemented with a 50%/50% (dry matter [DM], w/w) mixture of dried orange pellet and fresh orange peel to achieve a final concentration (DM, basis) of 0%, 10%, or 20% orange product (OP) for 10 days before inoculation with Salmonella Typhimurium. Sheep were experimentally inoculated with 1010 colony forming units Salmonella Typhimurium, and fecal samples were collected every 24 h after inoculation. Sheep were humanely euthanized at 96 h after oral Salmonella inoculation. Populations of inoculated Salmonella Typhimurium were numerically reduced by OP treatment throughout the gastrointestinal tract, and this reduction only reached significant levels in the cecum (p<0.05) of sheep fed 10% OP diets. Apparent palatability issues decreased the consumption of OP in sheep fed 20% OP to intake levels below that of 10% OP (approximately 7% dry matter intake [DMI]/d feed refusal), thereby reducing the potential effects of OP feeding at this higher level. Our results demonstrate that orange peel and pellets are environmentally friendly and low-cost products that can be used as a pre-harvest intervention as part of an integrated pathogen reduction scheme.
Introduction
A
Several phytochemicals have shown promise as antibacterials in a variety of applications (Cowan, 1999). Citrus essential oils have been part of the human diet for hundreds of years and their effects on bacterial growth and survival have been long studied (Dabbah et al., 1970; Neirotti et al., 1996; Viuda-Martos et al., 2008). Citrus oils kill Psuedomonas, Salmonella, Escherichia coli, and Staphylococcus aureus (Dabbah et al., 1970; Kim et al., 1995); recently, our research group demonstrated that the addition of >1% orange peel and pulp reduced populations of E. coli O157:H7 and Salmonella Typhimurium in mixed ruminal fluid fermentations in the laboratory (Callaway et al., 2008; Nannapaneni et al., 2008). Orange peel and pulp are byproduct feedstuffs that are commonly fed to cattle and have a good nutritive value for ruminants (6.9% crude protein [CP]; total digestible nutrients [TDN], 82%; net energy for maintenance [NEm], 1.9 Mcal/kg; net energy for growth [NEg], 1.3 Mcal/kg). Further studies have demonstrated that feeding orange peel and pulp reduced intestinal populations of diarrheagenic E. coli in weaned swine (Collier et al., 2010). The present study was designed to determine if using fresh and preserved orange peel and pulp as a dietary component could reduce gastrointestinal populations of Salmonella Typhimurium in an experimentally inoculated sheep model.
Materials and Methods
Bacterial cultures
S. enterica Typhimurium originally isolated from cattle and maintained in the Food and Feed Safety Research Unit culture collection was repeatedly grown by 10% (vol/vol) transfer in anoxic (85% N2, 10% CO2, 5% H2 atmosphere) Tryptic soy broth (TSB) medium at 37°C. This strain was selected for resistance to novobiocin and nalidixic acid (20 and 25 μg/mL, respectively) by repeated transfer and selection in the presence of sub-lethal concentrations of each antibiotic. This resistant phenotype was stable through multiple unselected transfers in batch culture and through repeated culture vessel turnovers in continuous culture (data not shown). Overnight cultures (1 L) were harvested by centrifugation (7500 g, 10 min) and cell pellets were re-suspended in TSB medium (150 mL total volume). Populations of Salmonella Typhimurium in these cell suspensions were determined to be approximately 2×109 colony forming units (CFU)/mL, by serial dilution and plating as described below.
Sheep, rations, and experimental design
All procedures in this study were approved by the Institutional Animal Care and Use Committee (IACUC protocol 05-001). Ramboullet/Suffolk sheep (average 60 kg body weight [BW]) were purchased from a commercial feedlot and were transported to the laboratory. Sheep were fed a commercial high grain diet composed of (dry matter [DM] basis): cracked corn 74.4%, soybean meal 9.2%, urea 0.7%, trace mineral salts 0.4%, and supplemented with coastal bermudagrass hay 15.3%. The diet was formulated according to National Research Council (NRC) recommendations and sheep were allowed ad libitum access to water.
Sheep were housed in environmentally controlled facilities and feces from each sheep was sampled on arrival and each subsequent day (n=7) during the dietary/facility adaptation period to verify that no organisms capable of growth on Brilliant Green agar (BGA) plates supplemented with novobiocin (20 μg/mL) and nalidixic acid (25 μg/mL) were present in the sheep. During this period no colonies grew on any of the BGA Nov/Nal plates.
Sheep (n=24) were randomly assigned to one of three treatment groups (n=8 sheep in each) that were fed diets supplemented with a 50/50% (DM basis, w/w) mixture of dried orange pellet and fresh orange peel to obtain final concentrations (DM basis, w/w replacement of concentrate portion of the ration) of 0%, 10%, or 20% orange product (OP; Texas Citrus Exchange, Mission, TX). Sheep were housed in enclosed facilities and were maintained on their respective diets for 10 days before the initiation of the study and throughout the entire study. Sheep were artificially inoculated with Salmonella Typhimurium (2×1010 CFU/sheep) via oral gavage (10 mL total volume per sheep) at 0 h. Every 24 h for 96 h after inoculation, fecal samples were collected via rectal grab from individual sheep and populations of inoculated Salmonella were determined.
Gastrointestinal sample collection
Sheep were humanely euthanized at 96 h. Contents and tissues from the rumen, distal end of the cecum, and the terminal rectum before the anal sphincter were aseptically collected upon necropsy. Digesta were serially diluted and plated as described below for quantitative enumeration of inoculated Salmonella Typhimurium populations. Tissues and digesta samples were enriched for inoculated Salmonella Typhimurium via a two-step process described below for detection. Gastrointestinal content pH was determined immediately upon return to the laboratory using a Corning 430 pH meter equipped with a calomel pH meter (Acton, MA). Intestinal contents were analyzed for volatile fatty acid concentrations (Corrier et al., 1990).
Bacterial enumeration
Ruminal, cecal, and rectal contents (10–20 g) as well as excreted feces were serially diluted (1 mL in10-fold increments) in phosphate buffered saline (pH 6.8) and directly plated on Brilliant Green Agar supplemented with novobiocin (20 μg/mL) and nalidixic acid (25 μg/mL; BGANN) plates. Colonies that grew on agar plates after 24 h incubation were directly counted (quantitative enumeration with a lower limit of detection of 101 CFU/mL). To qualitatively confirm the presence of inoculated Salmonella Typhimurium, intestinal contents and epithelial tissue samples as well as feces were enriched in tetrathionate broth at 39°C for 24 h followed by a secondary enrichment in Rappaport-Vasilladis broth at 42°C for 24 h with final plating on BGANN plates. Plates that contained typical Salmonella colonies after 24 h incubation were classified as positive for experimentally introduced Salmonella Typhimurium. Random colonies were picked during the course of the study and examined via growth on triple sugar iron agar and lysine iron agar (LIA) slants that were incubated at 37°C for 18 h to verify that the colonies growing on the BGANN plates were indeed Salmonella Typhimurium.
In vitro survival of inoculated Salmonella in feces
Feces from sheep (n=4 per treatment, n=12 total sheep) fed the three levels of orange peel (0%, 10%, and 20%) were collected, homogenized, and diluted (33%, w/v) into an anaerobic growth buffer that contained 1 g/L glucose (Callaway et al., 2005a). Fecal fermentations were incubated separately in sealed, crimped Balch tubes that were inoculated with 106 CFU/g Salmonella Typhimurium and were incubated at 39°C for 24 h. Fecal populations were determined by serial dilution and direct plating on BGANN at 24 h after inoculation.
Reagents and supplies
Unless otherwise noted, all media and agar were purchased from Difco Laboratories (Sparks, MD). Reagents and antibiotics were obtained from Sigma Chemical Co. (St. Louis, MO).
Statistics
Salmonella Typhimurium CFU/g were log10 transformed. Treatment groups were compared at each time point by the Mixed procedure of SAS (SAS Inst. Inc., Cary, NC). Time–treatment interactions were discounted due to the natural decay of Salmonella Typhimurium populations in this artificially inoculated model; therefore, only within time comparisons were performed for rumen, cecum, and rectal samples. Significance was determined at p<0.05.
Results
Apparent palatability issues decreased the consumption of OP in sheep fed 20% OP to intake levels below that of 10% OP (approximately 7% dry matter intake [DMI]/d feed refusal; data not shown); however, the group fed 10% consumed all OP daily. No colonies grew from TSB-enriched feces on the novobiocin/nalidixic acid-supplemented BGANN plates before the inoculation of the sheep with Salmonella Typhimurium, and sheep did not show any clinical signs of infection after inoculation. Salmonella populations in the feces of sheep fed 10% dried orange pulp were reduced numerically more than in controls or those fed 20% dried orange pulp at 96 h (Fig. 1).
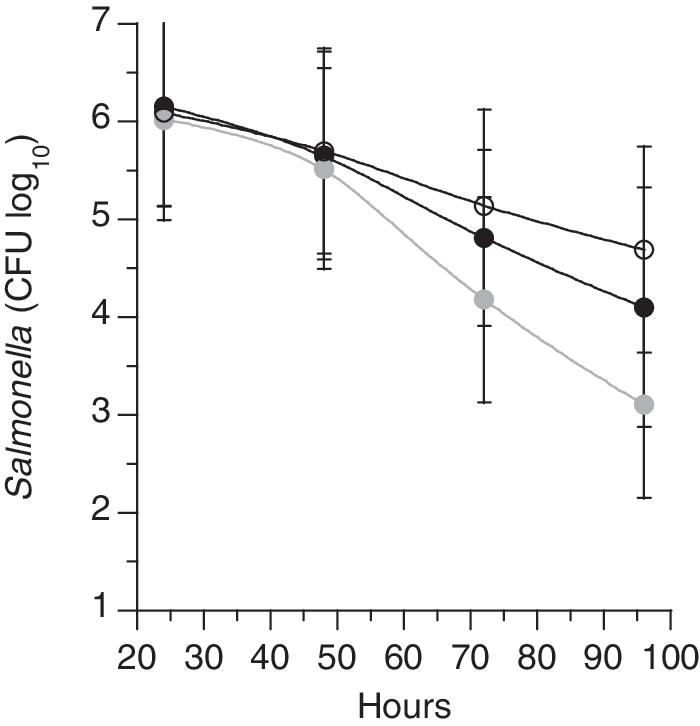
Fecal populations (CFU/g feces, log10) of Salmonella Typhimurium in sheep fed 0%, 10%, or 20% OP. Open circles indicate 0% OP, gray circles depict 10% OP diets, and black circles represent 20% OP. Error bars indicate standard deviations. CFU, colony forming unit; OP, orange product.
Ruminal Salmonella Typhimurium populations at necropsy at 96 h after inoculation were reduced by approximately 1 log10 CFU/g by 10% OP treatment, but this was not statistically significant (Fig. 2). Cecal populations of Salmonella Typhimurium were decreased by both levels of OP feeding, and were reduced 1.8 log10 CFU/g (p<0.05) by 10% OP feeding. Rectal Salmonella Typhimurium populations were not significantly impacted by either 10% or 20% OP feeding, but were lower numerically in the 10% OP fed sheep compared to 0 and 20% OP levels. Intestinal pH and volatile fatty acid concentrations were not different (p>0.05) in sheep fed OP or controls (data not shown).
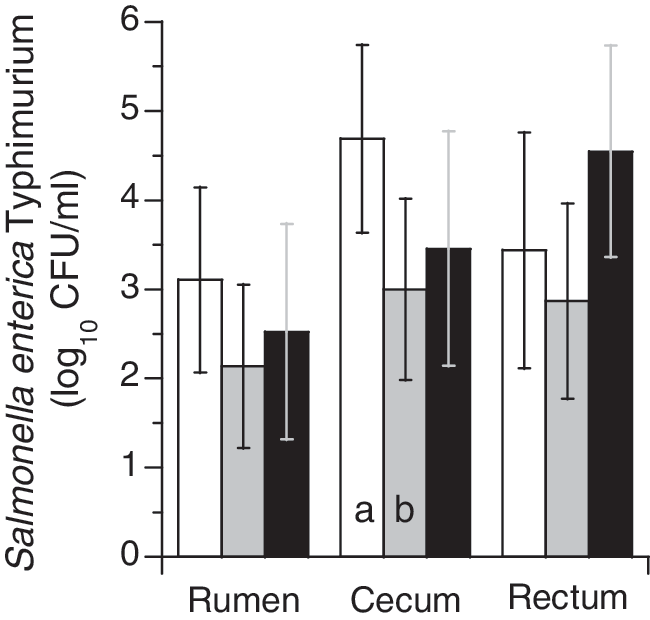
Intestinal populations (CFU/g feces, log10) of Salmonella Typhimurium in sheep fed 0%, 10%, or 20% OP. White bars represent 0% OP, gray bars 10% OP, and black bars 20% OP. Error bars indicate standard deviations. Different letters indicate statistical differences within a location (p<0.05).
When fecal fermentations from the sheep fed 0%, 10%, or 20% OP were inoculated with Salmonella in vitro, increasing levels of orange pulp inclusion in the diet reduced survival of Salmonella in the fecal in vitro fermentations (Fig. 3). Populations of Salmonella Typhimurium were reduced by approximately 0.75 (p<0.07) and 0.3 log10 CFU/g in 10% and 20% OP fed sheep, respectively.
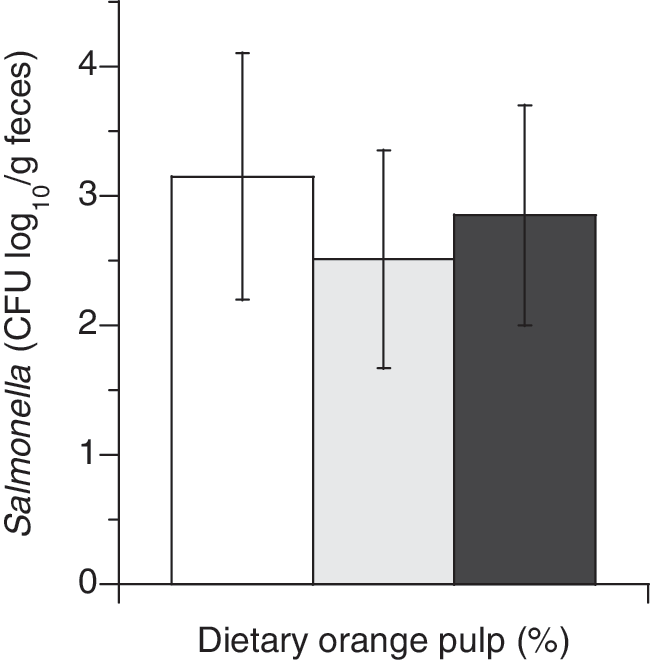
Salmonella populations (CFU/g feces, log10) after 24 h in vitro fermentations of feces from sheep fed 0%, 10%, or 20% OP diets. White bars represent 0% OP, gray bars 10% OP, and black bars 20% OP. Error bars indicate standard deviations.
Discussion
For more than a half a century, livestock producers have used antibiotics to enhance growth and to prevent disease in their herds (Gustafson and Bowen, 1997). However, in recent years, livestock producers have been under considerable pressure from consumer groups to reduce or eliminate the use of antimicrobials in livestock production due to a growing concern that this common production practice may lead to an increase in the number of antibiotic-resistant human pathogens (Corpet, 1996; Williams and Heymann, 1998). Due to these rising concerns, the European Union has banned the use of antibiotics as growth promotants and it is possible that U.S. policy-makers could follow suit. If antimicrobials are removed from livestock production, it will be imperative that U.S. producers have proven alternatives readily available to them if the livestock industry is to remain viable. Therefore, enhancing our knowledge in regard to how nutritional supplements may be used to improve the overall well-being and performance of the livestock, and provide nonantibiotic-dependent alternative management practices for the U.S. producer, is critical.
Salmonella is one of the most important foodborne pathogenic bacteria and is found in the intestinal tracts of poultry, swine, and ruminants (Davies et al., 1999; Callaway et al., 2005b; Vandeplas et al., 2010). While the implementation of pathogen reduction strategies in processing plants has been largely successful in reducing human illnesses compared historically (Koohmaraie et al., 2005), too many human foodborne illnesses still occur. As a result, there has been an increased focus on the development of preharvest intervention strategies that reduce pathogenic bacteria in food animals (Ransom et al., 2003; Sargeant et al., 2007; Vandeplas et al., 2010). If we can reduce pathogen populations in the live animal, then we lower the pathogen burden entering the processing facility, enhancing the effectiveness of current and future in-plant interventions. Furthermore, waste runoff from cattle facilities can contain foodborne pathogens, including Salmonella, which can contaminate water supplies (Manshadi et al., 2001; Johnson et al., 2003). Other routes of human exposures occur through direct and indirect animal contact (Oliver et al., 2008). Therefore, it is apparent that the application of the critical control point strategy to on-farm situations can produce intervention strategies that reduce human exposure to pathogens more than just within the processing plant (Oliver et al., 2008).
Orange peel and citrus pulp have excellent nutritional characteristics for cattle and have been fed for many years. They are low-cost ration ingredients that provide a good value to beef and dairy producers, and are typically included in cattle rations at levels of 5%–15% (Arthington et al., 2002). Earlier results from our research with citrus pulp and peel have indicated that these feedstuffs can impact populations of foodborne pathogenic bacteria in both pure and mixed ruminal fluid cultures in vitro (Callaway et al., 2008; Nannapaneni et al., 2008). In these studies populations of Salmonella were reduced by >3 log10 CFU/g by inclusion of up to 2% orange peel and pulp in the mixed ruminal microorganism fermentations (Callaway et al., 2008). In the present study, the inclusion of 10% OP decreased populations of Salmonella in the cecum more than 1 log10 CFU/g and reduced the rectal populations numerically, suggesting that even during a relatively short trial, orange peel can be a method to reduce foodborne pathogenic bacteria in food animals. Nevertheless, this reduction was only statistically significant in the cecum due to relatively small animal numbers coupled with population variability inherent in artificial infection models.
Plant-derived phenolic compounds containing reactive hydroxyl groups exhibit antimicrobial properties (Neirotti et al., 1996; Cowan, 1999); because of this activity, many naturally derived plant products (e.g., spices, fruit, or vegetable preparations/extracts), including citrus, have been used for centuries for the preservation and extension of the shelf life of foods (Dabbah et al., 1970). Citrus fruits contain a variety of compounds, including essential oils that exhibit activity against foodborne pathogens (Viuda-Martos et al., 2008; Friedly et al., 2009). In the present study the compound or oil fraction responsible for the observed antimicrobial activity was not determined; however, the active components of citrus oils were associated with limonene and the terpeneless fraction (Nannapaneni et al., 2008). Survival of inoculated Salmonella in in vitro fermentations was reduced numerically in fecal fermentations from sheep fed 10% OP compared to control sheep, suggesting that the antibacterial component of OP passes through the intestinal tract and kill pathogens in feces. This suggests the possibility that feces of animals fed orange pulp could exhibit anti-pathogen properties as the animals are shipped to slaughter and could reduce hide contamination with pathogens.
In the present study, increasing levels of OP over 10% negatively impacted intake, and therefore effectiveness of OP feeding. The intermediate level of actual intake by sheep fed 20% OP caused a reduction in the apparent impact of OP feeding; however, even at this lower intake level OP feeding reduced inoculated Salmonella populations numerically in the feces after 48 h of feeding and in the rumen and cecum upon necropsy. Thus, palatability issues apparently preclude intensive feeding of OP as a dietary “cleansing agent,” suggesting that feeding citrus products alone can reduce, but not eliminate carriage of foodborne pathogenic bacteria in the intestinal tract of food animals. This could indicate an important role for OP in feeding dairy or beef cattle in a long-term scenario, thus reducing the environmental impact or horizontal transmission, and in feeding to cattle before field days or open farms to prevent direct transmission to humans via contact.
It is important to note that the present study utilized an experimentally inoculated model to demonstrate the efficacy of a citrus peel feeding treatment within a relatively narrow window of Salmonella colonization. After experimental inoculation of ruminants, Salmonella intestinal populations decline in a manner that is not identical to the colonization of the intestinal tract. Furthermore, the populations inoculated in these sheep (1010 CFU total) is much higher than levels normally found in the gastrointestinal tract; therefore, products like citrus pulp and peel may have a greater impact in naturally infected animals. Based upon the present data, we conclude that the inclusion of orange peel and pulp in ruminant rations is a viable strategy to reduce Salmonella in the gastrointestinal tract before harvest. If we are to implement this pathogen intervention in the human food chain, further research needs to be performed to determine the most efficacious dosing strategies and levels of orange peel feeding. Orange peel and pulp feeding will not prevent all human foodborne illnesses; however, including citrus oils and products in the diet can be utilized in an integrated, multihurdle system aimed at reducing the passage of foodborne pathogens from farm to fork.
Footnotes
Acknowledgments
Portions of this research were supported by the U.S. Department of Agriculture and by beef and veal producers and importers through their $1-per-head checkoff, and were produced for the Cattlemen's Beef Board and state beef councils by the National Cattlemen's Beef Association.
Disclaimer
Proprietary or brand names are necessary to report factually on available data; however, the USDA neither guarantees nor warrants the standard of the product, and the use of the name by the USDA implies neither approval of the product, nor exclusion of others that may be suitable.
Disclosure Statement
No competing financial interests exist.