
Abstract
The prevalence of β-lactamase, 16S rRNA methylase genes, and plasmid-mediated fluoroquinolone-resistance (PMQR) determinants (qnrC and qnrD) was determined by polymerase chain reaction in fluoroquinolone-resistant Escherichia coli isolated from a chicken farm, a pig farm, and a hospital in Shandong, China in 2007. The bla TEM and bla CTX-M were the most prevalent β-lactamase genes in isolates from chickens (88.4%, 175/198 and 81.3%, 161/198) and hospitalized patients (87.8%, 122/139 and 69.1%, 96/139). The bla TEM was the most prevalent β-lactamase gene observed in isolates from pigs (98.5%, 135/137). The gene bla CMY-2 was also predominant among isolates from chickens (20.2%, 40/198). The bla LAP-1 gene was first detected in one strain from chickens and humans (pig farm workers) in China. Only one strain from hospitalized patients was found to possess bla SHV. The rmtB was the most prevalent 16S rRNA methylase gene detected in isolates from chickens (19.7%, 39/198) and hospitalized patients (15.8%, 22/139). To our knowledge, this is the first report of the detection of the qnrD gene in E. coli from chickens and pigs in China. The qnrC and bla KPC genes were not detected in any of the isolates. Results of southern hybridization revealed that PMQR determinants, β-lactamases, and 16S rRNA methylase genes were located on the same plasmid in E. coli strains derived from patients. Also, PMQR determinants and β-lactamase genes were localized on the same plasmid in an E. coli strain of animal origin. Results of conjugation experiments revealed that all of these plasmid-based resistance genes can be transferred by conjugation through horizontal transmission.
Introduction
Fluoroquinolones, β-lactams, and aminoglycosides have broad-spectrum antibacterial activity and have been widely used for the treatment of infection caused by a variety of bacterial pathogens. An increase in fluoroquinolone-resistant bacteria has been linked to the extensive clinical use of fluoroquinolones worldwide. Some hospitals reported rates of fluoroquinolone resistance above 50% among Enterobacteriaceae, which were once uniformly susceptible to fluoroquinolones (Wang et al., 2001). In Canada, resistance to ceftiofur observed in chicken-derived Escherichia coli from abattoir surveillance increased from 16% in 2002/2003 to 25% in 2004 (CIPARS, 2006). The yearly amikacin resistance rates of isolates from chickens in China varied from around 0.9% in 2002 to 6.8% in 2006 (Dai et al., 2008).
The plasmid-mediated fluoroquinolone-resistance (PMQR) determinants, β-lactamases, and/or 16S rRNA methylase genes often coexist and can also be associated with transposons/integrons, thereby increasing the potential enrichment of multidrug resistant bacteria by using multiple antimicrobial agents as well as by dissemination of resistance determinants among different bacterial species (Jiang et al., 2008; Liu et al., 2008; Kang et al., 2009; Shin et al., 2009).
The purpose of this study was to evaluate fluoroquinolone-resistant E. coli from different sources for β-lactamase, 16S rRNA methylase genes, and newly discovered PMQR determinants and the possibility of horizontal transmission of these genes.
Materials and Methods
The background of collection and identification of E. coli isolates
Fecal and farm environment samples were taken from one hospital, one pig farm, and one chicken farm in Shandong, China, during February 2007. All the samples were taken with disposable sterile swabs. The patients with diarrhea who had been admitted in the hospital and who had been treated with fluoroquinolone drugs and the farm workers from the chicken and pig farm voluntarily participated in this study, and were requested to collect their own fecal samples after defecation. The animals enrolled in this study were randomly selected from flocks and pens. Fecal samples from pigs and chickens were rectally collected from individual animals. Samples from the farm environment (including soil samples randomly taken from different places in the pig farm) and water (including pig and chicken drinking water) were collected.
A total of 597 E. coli isolates were confirmed using the API-20E system (BioMérieux). The minimum inhibitory concentration (MIC) of fluoroquinolones (including enrofloxacin, ciprofloxacin, and norfloxacin) for these isolates were obtained from Shandong Academy of Agricultural Sciences, Institute of Animal Husbandry and Veterinary Medicine. E. coli isolates (n=530) resistant to one or more fluoroquinolones (except 7 isolates from workers in the pig farm) were studied for the fluoroquinolone resistance genes.
The background of fluoroquinolone-resistant E. coli isolates harboring PMQR determinants
PMQR determinants [qnrA, qnrB, qnrS, aac(6′)-Ib-cr, and qepA] were analyzed in 530 fluoroquinolone-resistant E. coli isolates. The aac(6′)-Ib-cr was the most prevalent PMQR determinant and presented in isolates from all sources. The qnrS gene (63.2%, 108/171) was prevalent in isolates from the pig farm, whereas low prevalence was detected in bacteria recovered from hospitalized patients (2.2%, 3/139) and chickens (0.51%, 1/198). Only eight (5.8%, 8/139) isolates from hospitalized patients carried qepA. The qnrA and qnrB genes were not detected in any of the tested isolates (Xia et al., 2010).
Detection of β-lactamase, 16S rRNA methylase genes, and the newly discovered PMQR determinants
β-lactamase genes (bla TEM, bla CTX-M, bla SHV, bla LAP-1, bla KPC, and bla CMY-2), 16S rRNA methylase genes (armA and rmtB), and recently discovered PMQR determinants (qnrC and qnrD) in 530 fluoroquinolone-resistant E. coli isolates were analyzed by polymerase chain reaction (PCR). The bla TEM, bla CTX-M, bla SHV, bla LAP-1, qnrC, and qnrD genes were detected using previously described primers (Monstein et al., 2007; Poirel et al., 2007; Cavaco et al., 2009; Wang et al., 2009). Primers for the bla KPC, bla CMY-2, armA, and rmtB genes were designed in this study by Primer Premier 5.0. All primer sequences and the PCR conditions were shown in Table 1. The positive control, obtained from our laboratory, was sequenced and verified against a published sequence deposited in GenBank using the BLAST program.
Transfer of resistance and plasmid analysis
Fifteen isolates selected for conjugation experiments using the azide-resistant E. coli J53 as recipient, including eight isolates from patients, six isolates from pigs, and one isolate from chicken. They all carried at least two types of the following resistance gene combinations, such as PMQR determinants, β-lactamase, or 16S rRNA methylase genes. Transconjugants were selected on tryptone soy agar plates containing ciprofloxacin (0.1 μg/mL), azide (100 μg/mL), and ampicillin (100 μg/mL), respectively. Transconjugants harboring resistance genes were further confirmed by PCR. Plasmid DNA were extracted from donor strains and transconjugants using the QIAGEN Large-Construct Kit and then subjected to electrophoresis on a 0.8% Certified megabase gel (Bio-Rad Laboratory) without ethidium bromide staining. Plasmid sizes were estimated in comparison with plasmid size standards V517 (54, 5.6, 5.1, 3.9, 3.0, 2.7, and 2.1 kb), R1 (92 kb), and R27 (182 kb) using the computerized image analysis system (Alphaimager 2200 Pharmacia Biotech Co.).
Susceptibility testing of transconjugants and azide-resistant E. coli J53
MICs of antimicrobials were determined by the broth microdilution method according to the Clinical and Laboratory Standards Institute (CLSI, 2007, 2008). Antibiotics and the concentrations (μg/mL, in parentheses) tested were enrofloxacin (0.004–2.048), ciprofloxacin (0.002–1.024), norfloxacin (0.03–7.68), ampicillin (1–512), gentamicin (0.125–64), cefazolin (0.5–256), amoxicillin/clavulanic acid (1–512/2–1024), ceftiofur (0.25–128), amikacin (0.25–128), and apramycin (1–512). E. coli ATCC 25922 was used as the quality control strain.
Southern blot hybridization
Locations of the bla TEM, bla CTX-M, and rmtB genes on plasmids of eight human isolates containing the bla TEM and rmtB genes (including six isolates carrying the bla CTX-M gene) and four strains containing the bla TEM genes from pigs were determined by southern blot hybridization.
Locations of the bla TEM, bla CTX-M, rmtB, qepA, aac(6′)-Ib-cr, and qnrS genes on plasmids of transconjugants of the human strains containing the bla TEM, bla CTX-M, rmtB, qepA, and aac(6′)-Ib-cr genes and transconjugants of swine strains containing the bla TEM, qnrS and aac(6′)-Ib-cr genes were determined by southern blot hybridization.
Results
Prevalence of β-lactamase and 16S rRNA methylase genes
As shown in Table 2, bla TEM was the most prevalent β-lactamase gene in isolates from different sources. The bla CTX-M gene was prevalent among isolates from the chicken farm (80.1%, 178/220) and hospital (69.1%, 96/139), whereas low prevalence of bla CTX-M was detected in bacteria recovered from the pig farm (0.58%, 1/171). Next was the bla CMY-2 gene, which was predominantly present in isolates from the chicken farm (18.6%, 41/220). Only one isolate from chickens (0.51%, 1/198) and humans (pig farm workers) (0.64%, 1/156) was found to possess the bla LAP-1 gene, respectively. Only one isolate from patients (0.72%, 1/139) was found to possess the bla SHV gene. The rmtB was the most prevalent 16S rRNA methylase gene, which was detected in E. coli isolated from the chicken farm (18.6%, 41/220) and the hospital (15.8%, 22/139), and only one isolate from pigs (0.73%, 1/137) was found to possess the rmtB gene. The armA gene was detected only in isolates from chickens (1.0%, 2/198) and patients (4.3%, 6/139). In this study, the recently discovered qnrC and qnrD genes were detected. Only one isolate from chickens (0.51%, 1/198) and another isolate from pigs (0.73%, 1/137) were found to possess qnrD, respectively. The qnrC and bla KPC genes were not discovered in any of the isolates.
As shown in Table 3, among isolates from the chicken farm, the β-lactamases gene, 16S rRNA methylase gene, and PMQR determinants coexisted in 28 strains (12.7%). The coexistence of these three different kinds of resistance genes was the most frequently observed combination in isolates from the chicken farm. Further, this combination was also found in 22 strains (15.8%) from patients, but not found in strains from the pig farm.
Among the isolates from the chicken farm and hospital, 57 isolates (25.9%) and 23 isolates (16.5%) coharbored the bla CTX-M, bla TEM, and aac(6′)-Ib-cr genes, respectively, which was the most frequently observed combination among β-lactamase genes and PMQR determinants. Meanwhile, 42 strains (19.1%) and 40 strains (28.8%) coharbored the bla CTX-M and bla TEM genes, respectively, which was the most frequently observed combination among β-lactamase.
Among the isolates from the pig farm, 53 isolates (31.0%) coharbored the qnrS and bla TEM genes and 46 strains (26.9%) coharbored the qnrS, aac(6′)-Ib-cr, and bla TEM genes, which was the most frequently observed combination among PMQR determinants and β-lactamase genes.
Antimicrobial resistance of transconjugants and recipient to β-lactams, aminoglycosides, and fluoroquinolones
Eight strains (53.3%, 8/15) harboring multiple resistance genes were successfully conjugated with the recipient isolates. Conjugation was successfully preformed for the qnrS-, aac(6′)-Ib-cr-, and bla TEM-positive E. coli isolates P037, P066, P101, and P164; qnrS-, aac(6′)-Ib-cr-, and bla LAP-1-positive E. coli isolates C202; and qepA-, aac(6′)-Ib-cr-, rmtB-, bla CTX-M-, and bla TEM-positive E. coli isolates E36, E37, and E143, respectively.
The PMQR determinants-carrying plasmids were transferred into recipient strains. Increase in the MICs against three fluoroquinolones were observed in all transconjugants, a >2–64-fold increase in MICs for three fluoroquinolones from 0.064 to 2.056 mg/L (Table 4). MICs to other antimicrobials also increased in the transconjugants (Table 4), which suggested that the plasmids harboring β-lactamase and 16S rRNA methylase genes were transferred together.
Resistance breakpoints: enrofloxacin ≥2 μg/mL, ciprofloxacin ≥4 μg/mL, norfloxacin ≥8 μg/mL, ampicillin ≥32 μg/mL, gentamicin ≥16 μg/mL, cefazolin ≥32 μg/mL, amoxicillin/clavulanic acid ≥32/16 μg/mL and amikacin ≥64 μg/mL.
The names of the strains beginning with letter T indicate that they are transconjugants. The letters E, P, and C in the E. coli isolate number refers to patient, pig, and chicken, respectively. E36, E37, and their transconjugants harbored qepA, rmtB, bla TEM , bla CTX-M, and aac(6′)-Ib-cr genes, E143 and TE143 harbored qepA, rmtB, bla TEM, and aac(6′)-Ib-cr genes, P037, P066, P101, and P164 and their transconjugants harbored qnrS, aac(6′)-Ib-cr, and bla TEM genes, C202 and TC202 harbored qnrS, aac(6′)-Ib-cr, and bla LAP-1 genes.
Association of the genes with plasmids and antibiotic resistance
Southern hybridization revealed that bla TEM, parts of rmtB, and bla CTX-M genes were localized on plasmids and/or chromosome (Fig. 1). In our previous research, qnrS, qepA, and parts of aac(6′)-Ib-cr genes in isolates from different sources were shown to be on the same plasmid, respectively (Xia et al., 2010). The rmtB and bla TEM genes in isolates from patients were colocalized with qepA and parts of aac(6′)-Ib-cr genes on the same plasmids, respectively. The bla CTX-M gene in E2, E36, E37, and E130 was located on the other plasmid, respectively.
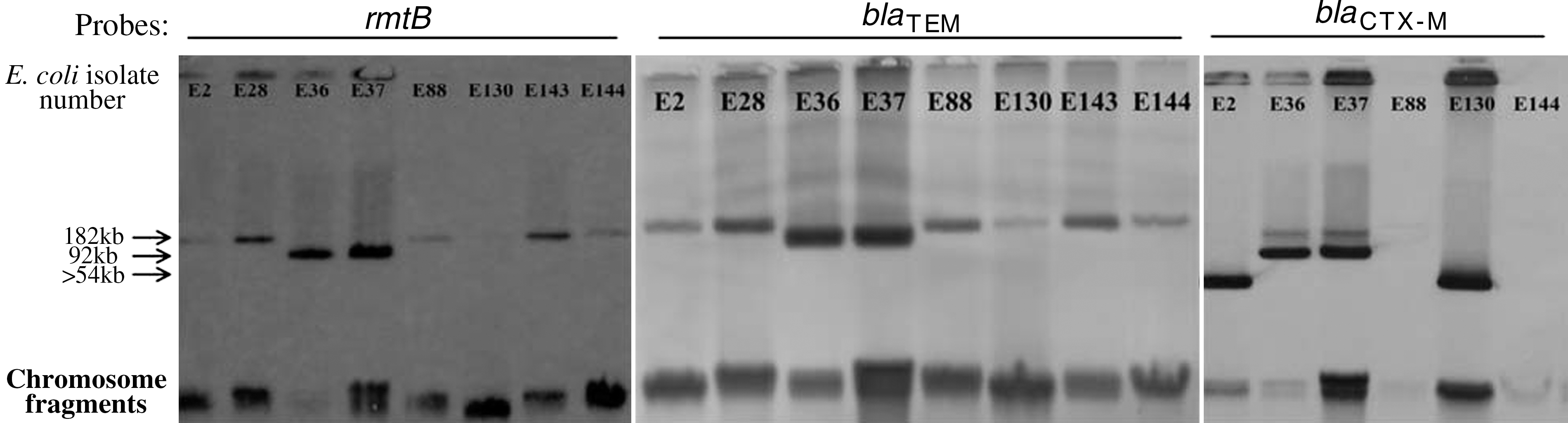
Southern hybridization of the extracted plasmid DNA from patients isolates with probes specific for bla TEM, bla CTX-M, and rmtB. The letter E in the Escherichia coli isolate number refers to patient.
The DNA probes of qnrS, aac(6′)-Ib-cr, qepA, rmtB, bla TEM, and bla CTX-M hybridized with plasmids extracted from the transconjugants (Figs. 2 and 3). In E36 and E37, the probes of bla CTX-M hybridized to one plasmid, with other genes [aac(6′)-Ib-cr, qepA, rmtB and bla TEM] not in the same plasmid, but in TE36 and TE37, the probes of bla CTX-M hybridized to the same plasmid with other genes. In P164, the probes of bla TEM hybridized to one plasmid, with other genes [aac(6′)-Ib-cr and qnrS] not in the same plasmid, but in TP164, the probes of bla TEM hybridized to the same plasmid with other genes. In P037, the two probes of qnrS and aac(6′)-Ib-cr hybridized to two different size plasmids. However, in TP037, the two probes hybridized to the same plasmid. Further, in P066, the three probes of qnrS, aac(6′)-Ib-cr, and bla TEM hybridized to three different size plasmids, respectively. However, in TP066, the three probes hybridized to the same plasmid.

Southern hybridization of the extracted plasmid DNA of transconjugants with probes specific for bla TEM, bla CTX-M, rmtB, qepA, and aac(6′)-Ib-cr. The names of the strains beginning with letter T indicate that they are transconjugants. The letter E in the E. coli isolate number refers to patient.
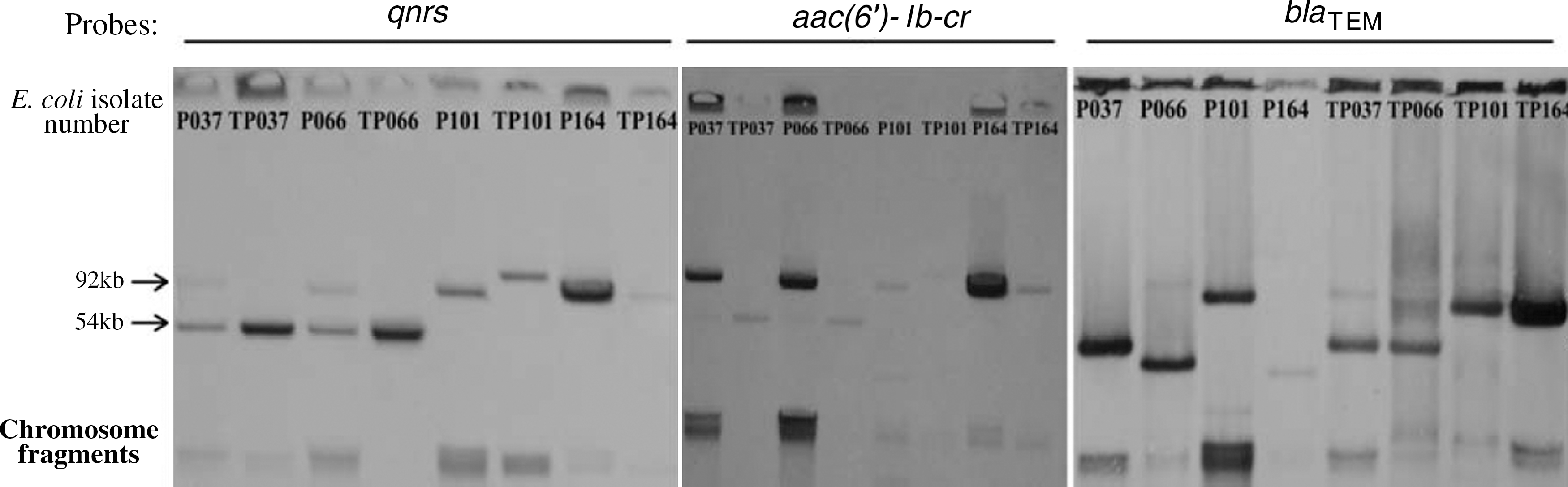
Southern hybridization of the extracted plasmid DNA of donor bacteria and transconjugants with probes specific for qnrS, aac(6′)-Ib-cr, and bla TEM. The names of the strains beginning with letter T indicate that they are transconjugants. The letter P in the E. coli isolate number refers to pig.
In P101, the three probes of qnrS, aac(6′)-Ib-cr, and bla TEM hybridized to one plasmid ∼90 kb. However, in TP101, the bla TEM probe hybridized to the plasmid of <90 kb, with other genes [aac(6′)-Ib-cr and qnrS] in another plasmid. Surprisingly, the band that hybridized to the bla TEM probe was invisible in electrophoresis.
Discussion
The bla TEM gene was more widespread than other β-lactamase genes detected among E. coli strains from different sources. The high detection rate of the bla TEM gene among isolates from different sources may be the primary responsibility for the high-ampicillin resistance of these strains. Some reports revealed that the narrow-spectrum TEM-1 or its derivatives were prevalent in ampicillin-resistant E. coli and Salmonella spp. from animals (Maidhof et al., 2002; Chen et al., 2004; Olesen et al., 2004).
The bla CTX-M was the second most prevalent gene in isolates from the hospital and the chicken farm, whereas this gene had low prevalence in pig farm isolates. In our previous study, compared with the chicken farm and hospital, resistance rates of isolates to ceftiofur and cefazolin from a pig farm were lower (Xia et al., 2010). The high-detection rate of the bla CTX-M gene might have the primary responsibility of increasing the resistance rates of isolates from the chicken farm and hospital to ceftiofur and cefazolin. The CTX-M enzymes are also found to coexist with other β-lactamases, particularly TEM-1 or its derivatives (Bonnet, 2004). Some reports revealed that the presence of more than one β-lactamase would raise the β-lactam resistance level and likely expand resistance to a broader range of β-lactams (Briñas et al., 2003; Teale et al., 2005), and this may potentially result in greater risks for failure of β-lactam therapy.
Previous reports revealed that the bla CMY-2 gene was prevalent in extended-spectrum cephalosporin-resistant E. coli and Salmonella spp. of animal origin (Allen and Poppe, 2002; Alcaine et al., 2005). Based on resistance patterns, the frequently occurring cefoxitin/ceftiofur resistant isolates of E. coli and Salmonella around the world were likely associated with the production of CMY enzyme(s) (Li et al., 2007). In this study, the bla CMY-2 gene was predominantly present in isolates from chickens, and the resistance to β-lactams among chicken isolates was the highest among the chicken farm, pig farm, and hospital isolates (Xia et al., 2010).
The bla SHV gene was only detected in one isolate from hospitalized patients. The reason might be that the SHV-type ESBLs were mainly detected in Klebsiella pneumoniae (David and Robert, 2005), or the SHV enzymes may have been replaced by CTX-M enzymes (Falagas and Karageorgopoulos, 2009). In this study, the bla LAP-1 gene was found to coexist with qnrS and aac(6′)-Ib-cr genes in one isolate from chicken, and with the bla CMY-2 gene in one isolate from a human (pig farm worker). Poirel et al. (2007) reported that the bla LAP-1 gene along with the qnrS1 gene was identified in Enterobacter cloacae isolates from France and Vietnam. The bla KPC gene was not detected in any of the isolates. These results show that carbapenems might be the most reliable therapeutic agents in the future.
The 16S rRNA methylase gene rmtB was more prevalent in bacteria isolated from the chicken farm and hospital than armA. 16S rRNA methylase-mediated high-level resistance to aminoglycosides has been recently reported in clinical isolates of Gram-negative bacilli from several countries (Yamane et al., 2005; Doi et al., 2007; Yu et al., 2007; Zong et al., 2008).
In this study, the qnrD gene was first reported in two strains, one from chickens and the other one from pigs. The qnrD gene was found to coexist with bla TEM, bla CTX-M, and bla CMY-2 genes in one isolate from chickens, and with the bla TEM gene in one isolate from pigs. Cavaco et al. (2009) reported that the qnrD gene was located on a small plasmid of about 4 kb in Salmonella enterica serovar Kentucky and Bovismorbificans strains of human origin. To clarify whether the qnrD gene from different sources was located in one plasmid of the same size and conclude the similarity of the plasmids, further studies need to be carried out.
The coharboring of PMQR determinants and β-lactamase genes was common among isolates in this study. The coharboring of aac(6′)-Ib-cr, bla TEM, and bla CTX-M was present in isolates from the hospital and the chicken farm. The coharboring of qnrS and bla TEM, and the coharboring of aac(6′)-Ib-cr, qnrS, and bla TEM were mainly present in isolates from the pig farm. A few of the PMQR, β-lactamase and 16S rRNA methylase genes coharboring isolates were present in the isolates from the hospital and the chicken farm. No isolate coharboring PMQR, β-lactamase, and 16S rRNA methylase genes was found among the isolates from the pig farm.
Results of southern hybridization revealed that PMQR determinants, β-lactamase, and/or 16S rRNA methylase genes were localized on plasmids and/or in the chromosome of donors and their transconjugants. Further, the transconjugants showed elevated MICs to fluoroquinolones compared with the initial recipients. The results revealed that fluoroquinolone resistance and the PMQR determinants were transferred into the recipient. We found that rmtB or armA genes were commonly identified with the coexistence of β-lactamase genes or PMQR determinants (see Fig. 2) in the same isolate of E. coli from the hospital and the chicken farm. Yu et al. (2009) reported that the plasmids harboring 16S rRNA methylase genes (rmtB and armA) usually coexisted in the same isolate and are co-transferred with ESBL genes on a self-transmissible conjugative plasmid to the recipients.
In this study, the qnrS gene transferred into the recipient E. coli J53, which was a different result from previous studies (Wu et al., 2009). Surprisingly, qnrS was localized on plasmids of different sizes in their donors, but it was localized on the same size plasmid in their transconjugants. This phenomenon may be attributed to the function of transposons/integrons.
These results and previous results indicate that the high prevalence of PMQR, β-lactamase, and 16S rRNA methylase genes in E. coli of different sources may be mainly attributable to horizontal and colony transfer of these resistance determinants. Plasmids may play an important role in transfer of resistance genes. Although the exact mechanism of multidrug resistance dissemination remains to be determined, transfer of plasmid-borne resistance genes, especially multiple resistance genes residing in the same plasmid, is likely to play an important role in the dissemination of resistance genes. In this study, it was common to find that multiresistance genes cohabit in a single isolate, or that different resistance genes may reside in one plasmid. In future studies, we will characterize the location and function of the PMQR determinants, β-lactamase genes, and 16S rRNA methylase genes. Moreover, flanking regions of the three type genes will be sequenced to determine common transfer elements.
Footnotes
Acknowledgments
The authors are grateful to Dr. Ming-Gui Wang (Institute of Antibiotics, Huashan Hospital, Fudan University, Shanghai, China) and professor Jacoby GA for kindly providing the reference strains azide-resistant E. coli J53, and E. coli V517, E. coli R1 and E. coli R27.
Funding
This study was supported by the Program for Chang Jiang Scholars and the Innovative Research Team at the University of China (No. IRT0866) and a grant from the Food Safety Key Technology Project of the China National Science and Technology Pillar Program in the eleventh 5-year plan period (2006 BAK02A03).
Disclosure Statement
No competing financial interests exist.