
Abstract
Pomegranate juice (PJ) and pomegranate polyphenolic extracts (PP) have antiviral effects against HIV-1, influenza, herpes, and poxviruses, and we recently demonstrated their effect against human noroviral surrogates. In the present study, the time-dependent effects of two commercial brands of PJ and PP at two concentrations (2 and 4 mg/mL) on the infectivity of foodborne viral surrogates (feline calicivirus FCV-F9, murine norovirus MNV-1, and MS2 bacteriophage) at room temperature for up to 1 h were evaluated. Each virus at ∼5 log10 plaque-forming units (PFU)/mL was mixed with equal volumes of PJ, or PP at 4 or 8 mg/mL, and incubated for 0, 10, 20, 30, 45, and 60 min at room temperature. Viral titers after each treatment were determined by standardized plaque assays and compared with untreated controls. Virus titer reduction by PJ and PP was found to be a rather rapid process, with ≥50% of titer reduction occurring within the first 20 min of treatment for all three tested viruses. Within the first 20 min, titer reductions of 3.12, 0.79, and 0.23 log10 PFU/mL for FCV-F9, MNV-1, and MS2, respectively, were obtained using PJ. FCV-F9, MNV-1, and MS2 titers were reduced by 4.02, 0.68, and 0.18 log10 PFU/mL with 2 mg/mL PP and 5.09, 1.14, and 0.19 log10 PFU/mL with 4 mg/mL PP, respectively, after 20 min. The mechanism of viral reduction by PJ and PP needs to be elucidated and clinical trials should be undertaken before recommending for therapeutic or preventive purposes.
Introduction
Recently, natural antimicrobials such as plant extracts have become increasingly popular options for viral reduction (Nair et al., 1999, 2002; Schlesinger et al., 2003; Weiss et al., 2005; Lipson et al., 2007a; Su et al., 2010a). The advantages for the use of natural plant antimicrobials include the absence of reported/observed toxic effects along with associated health benefits such as antioxidant, anticancer, anti-inflammatory, and antimicrobial properties (Prashanth et al., 2001; Negi and Jayaprakasha, 2003; Braga et al., 2005; Neurath et al., 2005; Pantuck et al., 2006; Miguel et al., 2007; Reddy et al., 2007; Haidari et al., 2009; Kasimsetty et al., 2009; Lee et al., 2010). For example, cranberry extracts were reported to be effective in reducing titers of influenza virus, waterborne rotavirus SA-11, bovine reovirus, feline calicivirus, and murine norovirus (Weiss et al., 2005; Lipson et al., 2007a, 2007b; Su et al., 2010a). Grape seed extract has been shown to inhibit the replication and expression of human immunodeficiency virus (HIV-1) (Nair et al., 1999, 2002). Pomegranate extracts have shown antiviral effects against influenza virus, HIV-1, and polioviruses (Konowalchuk and Speirs, 1976; Neurath et al., 2005; Haidari et al., 2009).
Recently, we demonstrated the antiviral effects of pomegranate juice (PJ) and a powder form of polyphenolic extract from fresh pomegranate fruit-POM™ (pomegranate polyphenolic extracts [PP]) against human norovirus surrogates and also the time dependence effect for only murine norovirus (MNV-1) using one concentration of PP (4 mg/mL) (Su et al., 2010b). As human noroviruses cannot be grown in cell culture (Duizer et al., 2004), culturable surrogates such as bacteriophage MS2 (Dawson et al., 2005), feline calicivirus (FCV-F9) (Steinmann, 2004), and MNV-1 (Wobus et al., 2004) are used in infectivity assays. Our previous study showed that PJ and PP had significant antiviral effects on the titer reduction of FCV-F9, MNV-1, and MS2 (Su et al., 2010b). To better understand the foodborne virus titer reduction kinetics of PJ and PP (at two concentrations: 2 and 4 mg/mL), the present study was undertaken. The main objectives were to (1) determine the titer reduction profiles of the three tested surrogates over 1 h treatment time at room temperature by PJ or PP at 2 or 4 mg/mL and (2) evaluate the pH effect of PJ on the titer reduction of these three surrogates.
Materials and Methods
Viruses, bacterial host, and cell lines
Crandell Reese Feline Kidney (CRFK) and mouse leukemic monocyte macrophage cell line (RAW) 264.7 cells were maintained in Dulbecco's modified Eagle's medium (DMEM)/Ham's F-12 (HyClone Laboratories, Logan, UT) containing 10% heat-inactivated fetal bovine serum (FBS; HyClone Laboratories) and 1×Anti-Anti (Antibiotic-Antimycotic; Invitrogen, Grand Island, NY) at 37°C in an atmosphere containing 5% CO2. Escherichia coli B-15597 was grown in 3% trypticase soy broth (TSB) at 37°C. Viral stocks of FCV-F9 and MNV-1 were prepared by infecting confluent CRFK and RAW 264.7 cells, respectively, and MS2 bacteriophage stock was prepared by infecting a 6 h culture of E. coli B-15597 at 37°C for ∼16 h; and viruses were harvested by centrifugation and filtration through 0.22-μm filters as described earlier (Su et al., 2009).
Antiviral effects of PJ and PP solutions
Two commercial brands of PJ, denoted as PJ1 and PJ2 (pH 3.4), were purchased from local grocery stores. PP was a gift from POM Wonderful (Los Angeles, CA). PJ and PP solutions at concentrations of 4 or 8 mg/mL were filter-sterilized before use (Su et al., 2010b). The pH of PJ was adjusted to pH 7.0 before use for virus treatment. Treatments were performed by aseptically mixing equal volume of each virus with PJ, PP, or water to obtain titers of ∼5 log10 plaque-forming units (PFU)/mL and incubated at room temperature for 0, 10, 20, 30, 45, and 60 min, respectively. The pH of the PP–virus mixture was determined to be 6.0 and 6.5 for 4 mg/mL and 2 mg/mL PP, respectively. After incubation, treatments of FCV-F9 and MNV-1 were neutralized by 10-fold dilutions in DMEM containing 10% FBS, and MS2 treatment was neutralized by 10-fold dilution in 3% TSB containing 3% beef extract. Plaque assays for evaluating the infectivity of the viruses were carried out in duplicates as described earlier (D'Souza and Su, 2010). All PJ and PP treatments were performed twice in independent experiments and the data were statistically analyzed.
pH effects of PJ
To study the pH effect of PJ, PJ was titrated with 1 M NaOH to pH 4.0, 5.0, 6.0, 7.0, and 8.0. Punicalagin, a high-molecular-weight, water-soluble polyphenol that accounts for >50% of the potent antioxidant activity of PJ (Gil et al., 2000), is reported to be comparatively stable from pH 4.0 to 9.0 based on absorbance values (Lu et al., 2011). Therefore, PJ at these pH values was mixed with equal volumes of MNV-1 or MS2 at a titer of ∼5 log10 PFU/mL or of FCV-F9 at a titer of ∼7 log10 PFU/mL (FCV-F9 at a titer of ∼5 log10 PFU/mL was fully inactivated after 1 h incubation with PJ at pH 3.4 to 8.0; thus, FCV-F9 at a higher titer of ∼7 log10 PFU/mL was used to study the pH effect) and incubated at room temperature for 1 h, and infectivity was determined using standardized plaque assays.
Infectious plaque assays
MNV-1 and FCV-F9 plaque assays using confluent RAW 264.7 and CRFK cells, respectively, in six-well tissue culture plates were carried out as described earlier (Su et al., 2010b).
MS2 plaque assays were conducted using E. coli B-15597 host as previously described (Bae and Schwab, 2008). The viral titer was calculated by multiplying plaque counts by the dilution factors. Titer reduction was calculated by subtracting the viral titer of treatments from the water control.
Statistical analysis
Results from the two replicate treatments and controls were analyzed using analysis of variance with SAS software (version 9.2, SAS Institute, Cary, NC) and Tukey's test.
Results
Time-dependent effects of PJ and PP on titer reduction of FCV-F9, MNV-1, and MS2
The pH of PJ was found to be 3.4 and was therefore adjusted to pH 7.0 before use to eliminate any effect associated with acidic pH. Incubation of equal volumes of PJ or PP with initial viral titers of ∼5 log10 PFU/mL was found to reduce the titers of FCV-F9, MNV-1, and MS2 over time and is shown in Figure. 1.

Changes in viral titers of FCV-F9
For FCV-F9, the titer reduction curves by PJ1 and PP were found to be different (Fig. 1a). Titer reductions of 1.39, 1.99, and 2.42 log10 PFU/mL for FCV-F9 were obtained upon immediate mixing with PJ1, 2 mg/mL PP, and 4 mg/mL PP, respectively. FCV-F9 titers continued to drop by 1.73, 2.03, and 2.67 log10 PFU/mL within the next 20 min of incubation with PJ1, 2 mg/mL PP, and 4 mg/mL PP, respectively. FCV-F9 titers after PJ1 treatment reached a plateau after 10 min treatment, whereas 2 mg/mL PP and 4 mg/mL PP decreased the FCV-F9 titer to undetectable levels within 30 and 20 min of incubation, respectively, at room temperature.
The titer reduction curves of MNV-1 by PJ1, 2 mg/mL PP, and 4 mg/mL PP followed similar trends to each other, but 4 mg/mL PP had a slightly greater effect than PJ1 and 2 mg/mL PP (p<0.05; Fig. 1b). Compared with FCV-F9, very little reduction in MNV-1 titer was observed upon immediate mixing with PJ1 or PP. MNV-1 titer was reduced by 0.79, 0.68, and 1.14 log10 PFU/mL by PJ1, 2 mg/mL PP, and 4 mg/mL PP treatment, respectively, after 20 min. A total reduction of 1.03, 1.04, and 1.40 log10 PFU/mL in MNV-1 titer was obtained after treatment with PJ1, 2 mg/mL PP, and 4 mg/mL PP, respectively, for 1 h.
The titer reduction curves of bacteriophage MS2 by PJ1, 2 mg/mL PP, and 4 mg/mL PP are shown in Fig. 1c. A reduction of 0.08, 0.05, and 0.12 log10 PFU/mL in titers was observed upon immediate mixing of MS2 with PJ1, 2 mg/mL PP, and 4 mg/mL PP, respectively. Approximately 0.23, 0.18, and 0.19 log10 PFU/mL reduction in MS2 titer was obtained within 20 min treatment with PJ1, 2 mg/mL PP, and 4 mg/mL PP, respectively. MS2 titers continued to drop only slightly by 0.11, 0.21, and 0.23 log10 PFU/mL, respectively, within the next 40 min incubation, resulting in a total MS2 titer reduction of barely 0.34, 0.39, and 0.42 log10 PFU/mL for PJ1, 2 mg/mL PP, and 4 mg/mL PP treatments. Thus, reduction in MS2 titers after treatment with PJ1 or PP was found to be minimal compared with MNV-1 and FCV-F9.
The reduction of FCV-F9, MNV-1, and MS2 titers over 1 h incubation time is summarized in Table 1. The common characteristics of the three tested viruses are that ≥50% of the titer reduction was achieved within the first 20 min of treatment. The reduction rates of the three viruses followed the order of FCV-F9>MNV-1>MS2 for both PJ1 and PP treatments. For comparison, PJ1 contains ∼3.6 mg/mL polyphenols (Aviram et al., 2008); therefore, the mix of PJ1 and virus contains ∼1.8 mg/mL PP. In this study, PP at the final concentration of 2 mg/mL should give comparable titer reduction as commercial PJs. From Fig. 1, the inactivation curves of MNV-1 and MS2 by PJ1 and 2 mg/mL PP are similar, but 2 mg/mL PP has greater effect on FCV-F9 than PJ1.
Within each row, different superscript letters denote significant differences between treatments (p<0.05). PJ1 and PJ2 are pomegranate juice from two different manufacturers. Pomegranate polyphenol extracts (PP) were obtained from POM Wonderful (Los Angeles, CA).
PFU, plaque-forming units.
This study also compared the antiviral effects of PJ from two different commercial sources with similar expiration dates (named as PJ1 and PJ2, respectively, as shown in Table 1). There appeared to be only slight differences between the antiviral effects of the two brands of PJ upon mixing with FCV-F9 and MNV-1. However, the total titer reduction by PJ1 and PJ2 after 1 h incubation at room temperature was not significantly different (p<0.05) for the three tested viruses. During this study, we did notice that the antiviral effect of PJ decreased with storage time, which could be potentially due to the fact that punicalagin, the bioactive compound in PJ, possibly degraded to ellagic acid during storage based on earlier reports (Gonzalez-Molina et al., 2009).
The effect of pH on the antiviral activity of PJ against FCV-F9, MNV-1, and MS2
To study the pH effect of PJ on its antiviral activity, PJ was titrated to pH over the range of 3.4–8.0 before mixing with equal volumes of FCV-F9, MNV-1, and MS2. The obtained titer reductions after 1 h incubation are shown in Figure 2.
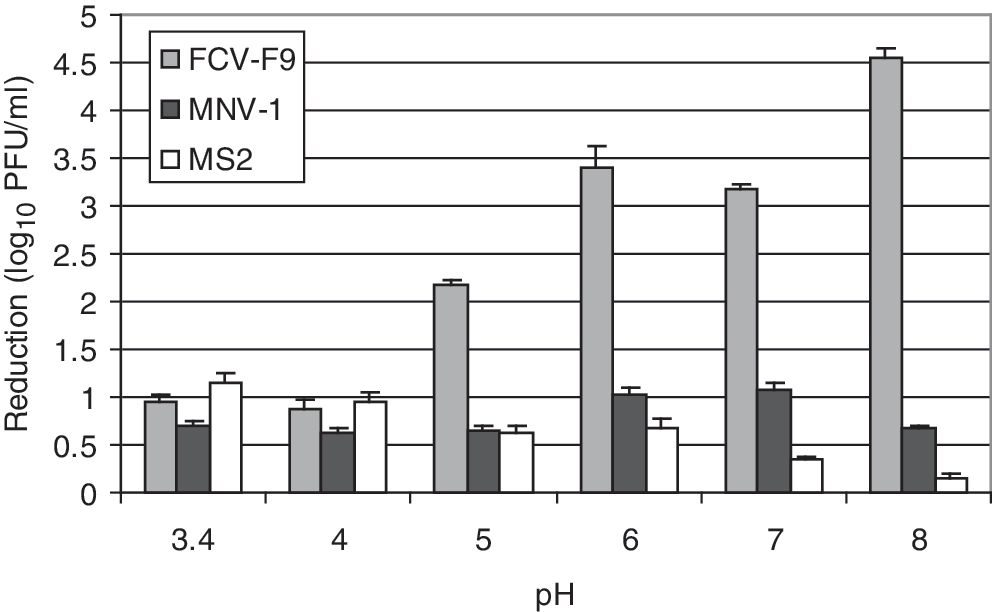
The effect of PJ with different pH values on the titer reduction of FCV-F9, MNV-1, and MS2 after 1 h incubation at room temperature. The pH of PJ (natural pH of 3.4) was adjusted to 4.0, 5.0, 6.0, 7.0, and 8.0 with 1 M NaOH before mixing with each virus. The titer of FCV-F9, MNV-1, and MS2 after 1 h treatment was evaluated using standardized plaque assays.
Among the three tested viruses, FCV-F9 was the most sensitive to changes in the juice pH. At pH 3.4, 4.0, and 5.0, FCV-F9 titer was reduced by 1–2 log10 PFU/mL. As the pH of PJ increased, the titer reduction also increased. FCV-F9 was reduced by 3.4, 3.2, and 4.6 log10 PFU/mL after PJ treatment at pH 6.0, 7.0, and 8.0, respectively.
The pH effect of PJ on MS2 reduction differed from that of FCV-F9. As the pH of the PJ increased, the titer reduction of MS2 decreased, wherein ∼1 log10 PFU/mL reduction was obtained with PJ at pH 3.4, but only 0.15 log10 PFU/mL reduction was achieved with PJ at pH 8.0.
For MNV-1, ∼1 log10 PFU/mL titer reduction was achieved by PJ treatment at pH 6.0 and 7.0. At the other pH values, MNV-1 titer reductions were ∼0.6 log10 PFU/mL. Thus, MNV-1 was less sensitive to pH changes in PJ than FCV-F9 and MS2.
Discussion
To eliminate the effect of pH, PJ was neutralized before use for virus treatment. Yet, PJ at neutral pH still resulted in reduction of ∼3 log10 PFU/mL for FCV-F9, ∼1 log10 PFU/mL for MNV-1, and ∼0.3 log10 PFU/mL for MS2. These results show that the acidic pH is not the only factor that contributes to the antiviral effect of PJ and that PP or other components in PJ also played a role in viral reduction of FCV-F9, MNV-1, and MS2. Moreover, PP at 2 mg/mL (the same PP concentration as in PJ) caused comparable titer reduction in all three viruses as PJ at pH 7.0. This again indicates that PP alone can have antiviral effects against FCV-F9, MNV-1, and MS2.
The antiviral effects of PP have been reported against HIV-1, influenza viruses, and poliovirus (Konowalchuk and Speirs, 1976; Neurath et al., 2005; Haidari et al., 2009; Sundararajan et al., 2010). Sundararajan et al. (2010) showed that PP at 0.2 mg/mL decreased influenza viral titers from 4 to 5 log10 PFU/mL to an undetectable level after 5 min at room temperature. Pomegranate extracts at 2 μg/mL was shown to reduce influenza A by ∼1.5 log10 PFU/mL after incubation at 4°C for 1 h as a result of replication inhibition (Haidari et al., 2009). Incubation of HIV-1 with PJ at 1:40 dilution for 1 h at 37°C was shown to result in >1 log10 PFU/mL reduction through inhibition of gp120-CD4 binding (Neurath et al., 2005). Konowalchuk and Speirs (1976) also found that pomegranate extracts could cause >2 log10 PFU/mL poliovirus reduction after mixing at 4°C for 24 h, but the mechanism of action was unknown.
In this study, FCV-F9 was shown to be more sensitive to the pH of PJ than MNV-1. This observation was consistent with earlier findings: Cannon et al. (2006) reported that by mixing viruses with 100 mM citrate buffer at low pH values of 2, 3, and 4, MNV-1 titer was reduced by 0.6, 0.6, and 0.5 log10 PFU/mL, respectively, whereas the FCV titer was reduced by 4.4, 3.7, and 2.3 log10 PFU/mL, respectively. Higher reduction at pH 8 could be attributed to the possible increase in availability of the PP at this pH or due to optimal activity of PP at near neutral pH values for FCV-F9 and MNV-1, again indicating that the majority of the effect was related to the components of PJ and not pH alone.
For PJ or PP treatment, the reduction trend followed the order of FCV-F9>MNV-1>MS2, similar to the trend observed with CJ (Su et al., 2010a). For comparison to chemical inactivation, our previous studies with trisodium phosphate (TSP), house-hold bleach, and glutaraldehyde (D'Souza and Su, 2010) showed that with 1% TSP, titer reductions followed the order of MS2>FCV-F9>MNV-1. For 1% glutaraldehyde treatment, the titer reductions followed the order of FCV-F9>MNV-1>MS2, and for 10% bleach, it was FCV-F9=MS2>MNV-1 (D'Souza and Su, 2010). The variability in virus reduction by different compounds could be due to the differences in chemical composition and pH of the chemicals used.
Additionally, Bae and Schwab (2008) found that at 25°C the reduction rate followed the order of FCV>MS2>MNV-1 in environmental and ground water, with reduction trends of FCV>MNV-1>MS2 in surface water (Bae and Schwab, 2008). Thus, the reduction rate of the viruses varied with the chemical agent as well as the environmental conditions. When FCV-F9 and MNV-1 were compared, long-term resistance was found to be higher for MNV-1 than FCV-F9 at room temperature, and MNV-1 was consistently more resistant than FCV-F9 to basic and acidic pH and chemical treatments (Cannon et al., 2006; Bae and Schwab, 2008; Macinga et al., 2008; Su et al., 2009). FCV-F9 could be more sensitive to treatments than the other two viruses because of the possible ease of structural damage to the FCV-F9 capsid. However, our results showing reduction of MNV-1 titers by PJ and PP, even though to a relative small extent of ∼1 log, are still quite promising, considering that MNV-1 is highly resistant to most treatment processes, and remains persistent in the environment. Also, some researchers consider MNV-1 to be one of the most appropriate cultivable surrogates for human noroviruses (Sosnovtsev et al., 2006; Wobus et al., 2006).
PJ and its extracts have been used to extend the shelf life of chicken products, ground beef, and ground goat meat (Lucas and Were, 2009; Devatkal and Naveena, 2010; Kanatt et al., 2010); chicken meat shelf-life was extended by 2–3 weeks during chilled storage and sensory studies showed no effect on the sensory attributes of the treated products (Kanatt et al., 2010). Currently, there is a growing interest in using natural preservatives and eco-friendly products to enhance the safety of ready-to-eat foods. Pomegranate extracts possess a desirable characteristic flavor along with antioxidant and antimicrobial properties (Haslam, 1996; Aviram et al., 2002; Machado et al., 2002; Taguri et al., 2004; Cheng et al., 2005; Liu et al., 2007, 2009). Therefore, these natural extracts can be used as part of hurdle approaches along with food processing technologies to enhance the food safety of ready-to-eat products to meet FDA standards and eliminate low-level viruses that can be present in foods and cause infection. Also, as the PP effect appears to be rather rapid, postprocessing contamination of foods by viruses can be controlled along with added health benefits and desirable flavors. In addition, novel nanotechnology delivery mechanisms of PPs for rapid release in the gastrointestinal tract to obtain maximum benefit and bioavailability studies can be undertaken based on this study showing the time dependence effects over 1 h. These data will be also useful for potential pharmaceutical or nutraceutical applications, wherein release kinetics upon encapsulation can be studied. These natural compounds can also be incorporated into edible films in wraps as well as other food dressings. However, further in vivo studies and clinical trials are needed.
Conclusions
The present study demonstrated that PJ and PP at 2 or 4 mg/mL resulted in varied reduction rates in the titers of FCV-F9, MNV-1, and MS2 over 1 h. Commercial brands did not appear to have significant differences in titer reductions, but storage time could play a role in the level of reduction observed. Among the three viruses, FCV-F9 was the most susceptible, and MS2 was the most resistant to PJ and PP treatments. Regardless of the difference in titer reduction rates, ≥50% of the total reduction was achieved with 20 min of treatment. Thus, PJ and PP appear to be promising natural antivirals that are effective almost immediately upon contact with the foodborne viral surrogates. However, our results were obtained using in vitro studies with cell culture systems and pure viral stocks. Further studies are necessary to evaluate the effect of PJ and PP in complex food environments and also their effects in vivo. Clinical trials and FDA and regulatory approvals are required before they can be recommended or even suggested for routine therapeutic or preventive use.
Footnotes
Acknowledgments
Funding for this research was provided by the Tennessee Agricultural Experiment Station (UT-TEN-HATCH No. 00391
Disclaimer
The use of trade names in this article does not imply endorsement by the University of Tennessee or criticism of similar ones not mentioned.
Disclosure Statement
No competing financial interests exist.