
Abstract
In the present study, Listeria monocytogenes BCRC 14846 and Salmonella Typhimurium BCRC 10747 were subjected to acid adaptation at pH 5.5 at 37°C for 1 and 4 h, respectively. The viability of the acid-adapted cells of test organisms exposed to Clidox-S, a chlorine-containing disinfectant, and Quatricide, a quaternary ammonium compound, was examined and compared with that of the control cells at 25°C and 40°C. Results revealed that acid adaptation significantly enhanced the viability of L. monocytogenes and Salmonella Typhimurium exposed to the disinfectants under investigation. Both pathogens examined were more susceptible to Clidox-S and Quatricide at 40°C than at 25°C. Further, L. monocytogenes was more susceptible to Quatricide than Salmonella Typhimurium, whereas Salmonella Typhimurium was more susceptible to Clidox-S than L. monocytogenes.
Introduction
T
Along with the disinfection, acidification is one of the general practices employed to control the growth of microorganisms in food-processing plants and households. However, it was reported that microorganisms might develop enhanced resistance to acidic and other stress environments after exposure to a sublethal acidic condition (acid adaptation response) (Leyer and Johnson, 1993; Lou and Yousef, 1997; Leenanon and Drake, 2001; Tiwari et al., 2004; Chen et al., 2009a). The enhanced resistance of microorganisms induced by sublethal acid stress has important implications in food safety and warrants concern. It raises the possibility that the acid adaptation response of microorganisms may render the currently employed microbial control measures, which have been developed based on studies using unstressed microorganisms, inadequate to the preservation of food safety. So far, numerous investigators have studied the susceptibility of microorganisms to disinfectants (Taormina and Beuchat, 2001; Peng et al., 2002; Sutton et al., 2002; Kim et al., 2007). While reports concerning the effect of acid adaptation on their viabilities exposed to disinfectants are still relatively limited. In the present study, two foodborne pathogens—Listeria monocytogenes BCRC 14846 and Salmonella Typhimurium BCRC 10747—were used as the test organisms. Viability of the acid-adapted and nonadapted cells of these pathogens exposed to disinfectants such as Clidox-S and Quatricide at 25°C and 40°C was investigated.
Materials and Methods
Test organisms
In the present study, L. monocytogenes BCRC 14846 and Salmonella Typhimurium BCRC 10747 obtained from the Bioresources Collection and Research Center (Hsinchu, Taiwan) were used as the test organisms.
The physiological activity of Salmonella Typhimurium and L. monocytogenes was first activated by two successive transfers in Tryptic soy broth (TSB; Difco) and TSB+yeast extract (Difco) (TSBYE), respectively, at 37°C for 12 h. The activated cultures were then grown in TSB or TSBYE for 12 h to enter their late log phase and were used as the inocula to prepare the acid-adapted and control cells.
Disinfectant solution
In the present study, two disinfectants were examined: Clidox-S, a choline base disinfectant, and Quatricide, a QACs with n-alkyl dimethyl ethylbenzyl ammonium chloride and n-alkyl dimethyl benzyl ammonium chloride as the active ingredients (Pharmacal Research Lab Inc., 2010). Both disinfectants are products of Pharmacal Research Labs Inc. The parent solution of these disinfectants was first prepared according to the manufacturer's instructions. The Clidox-S solution was prepared by mixing Clidox-S, distilled water, and Clidox-S activator at a ratio of 1:8:1 (v:v:v). The Quatricide solution was prepared using two ounces of Quatricide mixed with 1 gallon of distilled water (w/v). Based on results obtained from the preliminary experiments, the test disinfectant solutions (0.2% Clidox-S, 0.2% and 0.5% Quatricide) were further prepared by mixing an adequate amount of the parent disinfectant solution with sterile RO water. The test disinfectant solutions showed a pH of 7.1–7.4 and remained unchanged during the experimental period.
Acid adaptation of L. monocytogenes and Salmonella Typhimurium
Cultures of test organisms were first centrifuged (3000× g) for 20 min at 37°C. The pellets were washed twice with phosphate buffer solution (PBS, 0.1 M, pH 7.0).
The method described by O'Driscoll et al. (1996) and Shen et al. (2007), respectively, was followed to prepare the acid-adapted cells of L. monocytogenes and Salmonella Typhimurium. Essentially, the washed cells mentioned above were suspended in PBS that had their pH adjusted to 5.5 with 6 N HCl and incubated at 37°C for 1 and 4 h, respectively, for acid-adapted L. monocytogenes and Salmonella Typhimurium. The washed cells suspended in PBS (pH 7.0) served as the nonadapted (control) cells.
The viability of L. monocytogenes and Salmonella Typhimurium exposed to disinfectants
To determine the viability of test organisms exposed to disinfectants, 1 mL of acid-adapted and control cell suspension of L. monocytogenes or Salmonella Typhimurium was inoculated into 50 mL of the diluted Clidox-S or Quatricide solution at a population of ca 106–7 cfu/mL. They were then incubated at 25°C or 40°C for for 50 min when exposed to Quatricide. On the other hand, viability of Salmonella Typhimurium and L. monocytogenes was examined by exposure to Clidox-S for 5 and 25 min, respectively. At specific exposure intervals, 1 mL of sample was withdrawn and combined with 1 mL of Dey-Engley neutralizing broth (Sigma) to neutralize components that might be lethal to the test organisms (Sutton et al., 1991).
The enumeration of viable L. monocytogenes and Salmonella Typhimurium
To determine the viable population of test organisms, samples were serially diluted in PBS and the viable counts were made by pour plating on Tryptic soy agar (TSA; Difco) and TSA+yeast extract (TSAYE) for Salmonella Typhimurium and L. monocytogenes, respectively, after 24 h of incubation at 37°C.
Statistical analysis
The mean value and standard deviation were calculated from the data obtained from three separate experiments. In each experiment, two samples were taken at the specific time intervals for the determination of the viability of the test organism. Data were analyzed using unpaired two-tailed Student's t test. Statistical significance was set at p<0.05.
Results and Discussion
Acid adaptation affects the viability of L. monocytogenes exposed to Clidox-S
Various microorganisms have been reported to show enhanced resistance to lethal stresses when subjected to a prior acid adaptation (Leyer and Johnson, 1993; O'Driscoll et al., 1996; Lou and Yousef, 1997; Tsai and Ingham, 1997; Leenanon and Drake, 2001; Chen et al., 2009b). In the present study, L. monocytogenes was subjected to acid adaptation at pH 5.5 for 1 h using the method described by O'Driscoll et al. (1996). They reported that acid adaptation significantly enhanced the resistance of L. monocytogenes when it was subsequently exposed to lethal stresses such as pH 3.5, 55°C, 2.5% NaCl, and 15% alcohol. Figure 1 shows the viability of the acid-adapted and nonadapted cells of L. monocytogenes during the exposure to Clidox-S (0.2%) for 25 min at 25°C and 40°C. It was found that the viability of the test organism, regardless of acid adaptation, decreased as the period of exposure to Clidox-S at 25°C was extended (Fig. 1A). However, the acid-adapted L. monocytogenes generally exhibited a higher viability with a lower population reduction than the control cells at similar exposure intervals. Population reduction was obtained by subtracting the final viable population from the initial population. For example, the viable population of the control L. monocytogenes decreased from ∼6.88 log cfu/mL at 0 time to 3.80 log cfu/mL with a population reduction of 3.08 log cfu/mL after 25 min of exposure to Clidox-S. Meanwhile, the acid-adapted cells reduced from 6.89 to 5.21 log cfu/mL and showed a significantly lower (p<0.05) population reduction of 1.68 log cfu/mL compared with that of the control cells. This suggested that acid adaptation enhanced the survival of L. monocytogenes exposed to Clidox-S at 25°C.
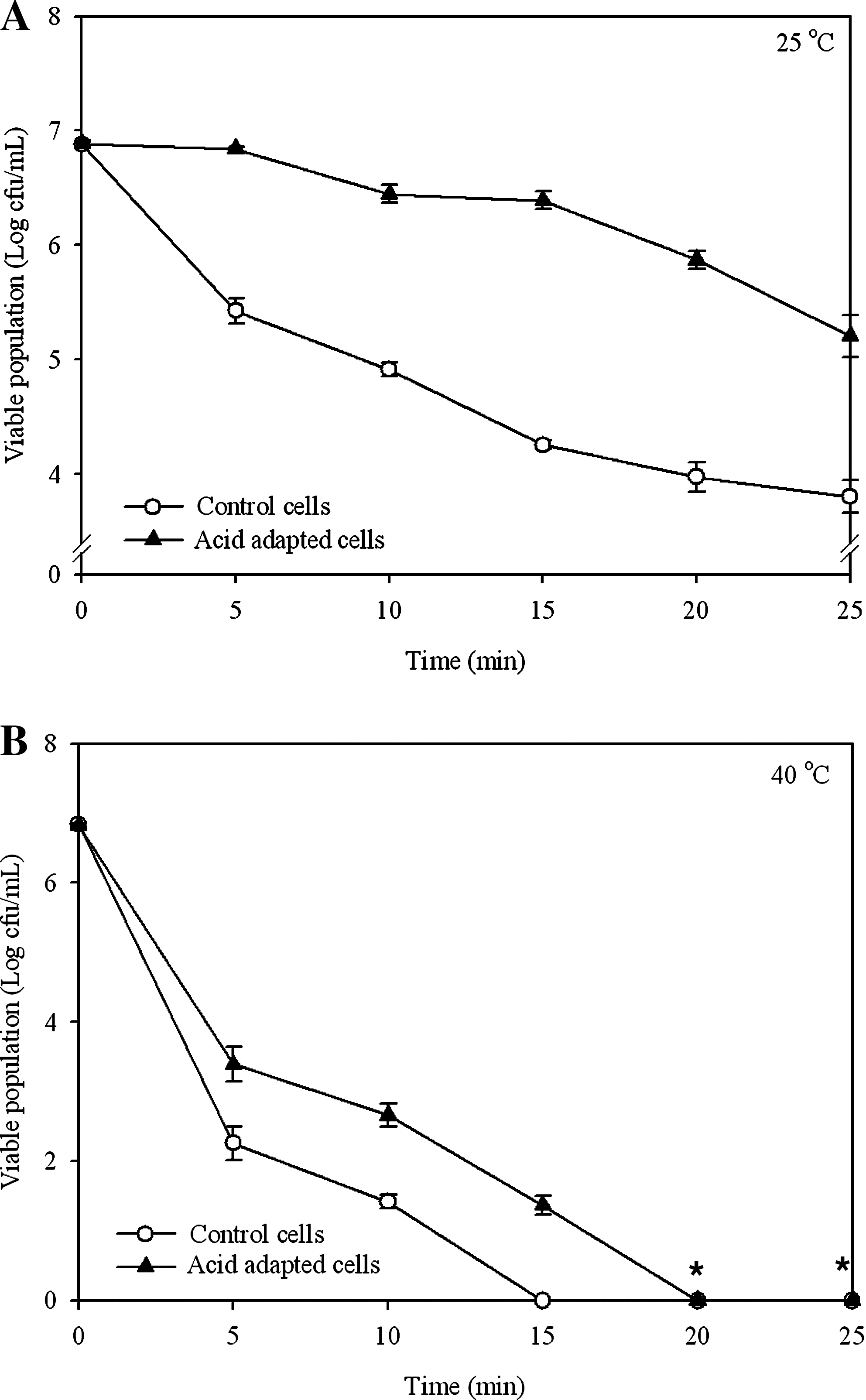
Viability of acid-adapted and control Listeria monocytogenes during exposure to Clidox-S (0.2%) at 25°C
Figure 1B shows the viability of acid-adapted and control L. monocytogenes after exposure to Clidox-S (0.2%) at 40°C. In general, a higher viability was noted with the acid-adapted cells than the controls. At 40°C, the acid-adapted and the control L. monocytogenes also showed a similar trend in the change of viability (Fig. 1B) as that noted at 25°C (Fig. 1A). Nevertheless, at 40°C, the rate of reduction in the viability of the test organism, regardless of acid adaptation, is greater than that noted at 25°C. These results further showed that L. monocytogenes, regardless of acid adaptation, is more susceptible to Clidox-S at higher temperature.
Acid adaptation affects the viability of L. monocytogenes exposed to Quatricide
As shown in Fig. 2A, with the extension of the exposure period to Quatricide (0.2%) at 25°C, the viable population of the acid-adapted and the control cells of L. monocytogenes was reduced. With an initial population of ∼6.0 log cfu/mL, the acid-adapted and the control cells reduced to 4.68 and 3.35 log cfu/mL, respectively, after 50 min of exposure when the acid-adapted L. monocytogenes showed a significantly lower (p<0.05) population reduction of 2.19 log cfu/mL than that (log 3.37 log cfu/mL) of the controls. This phenomenon was noted throughout the entire exposure period and implied that acid adaptation enabled L. monocytogenes to become less susceptible to Quatricide as it had with Clidox-S (Fig. 1A). Similar to observations on Clidoxide-S (Fig. 1A), greater antibacterial activity was exerted by Quatricide at 40°C than at 25°C.

Viability of acid-adapted and control L. monocytogenes during exposure to Quatricide (0.2%) at 25°C
Acid adaptation affects the viability of Salmonella Typhimurium exposed to Clidox-S
The prolonged survival of acid-adapted Salmonella Typhimurium in food systems that have both high acidity and that are kept at refrigerated temperature have been observed and have been attributed to the enhanced acid tolerance of the test organism (Leyer and Johnson, 1993; Tsai and Ingham, 1997; Shen et al., 2007). Additionally, Leyer and Johnson (1993) observed that acid adaptation increased tolerance of Salmonella Typhimurium toward various stresses, including heat, salt, an activated lactoperoxidase system, crystal violet, and polymyxin B. Moreover, they also reported that specific out membrane proteins were induced in the acid-adapted Salmonella Typhimurium and indicated that the mechanism of acid-induced cross-protection involved changes in cell surface properties in addition to the enhancement of intracellular pH homeostasis.
Figure 3 shows the viability of acid-adapted and control cells of Salmonella Typhimurium during exposure to Clidox-S (0.2%) at 25 and 40°C. The viable population of Salmonella Typhimurium, regardless of acid adaptation, decreased during the exposure period. In addition, acid adaptation also increased the resistance of Salmonella Typhimurium to Clidox-S. At 25°C and at the end of the 5 min of exposure period, the acid-adapted Salmonella Typhimurium showed a viable population of 4.43 log cfu/mL (Fig. 3A) with a population reduction of 2.45 log cfu/mL, which is significantly less (p<0.05) than that of 3.18 log cfu/mL observed with the control cells. At 40°C, no viable cells of the control Salmonella Typhimurium was detectable after 4 min of exposure to Clidox-S, whereas a longer exposure period of 5 min is required for Clidox-S to exert a similar effect on the acid-adapted cell (Fig. 3B). These results all demonstrated that Salmonella Typhimurium was more susceptible to Clidox-S at 40°C than at 25°C and acid adaptation reduced the susceptibility of Salmonella Typhimurium to Clidox-S. Additionally, it was found that control cells of Salmonella Typhimurium showed a viable population of 3.69 log cfu/mL with a population reduction of 3.18 log cfu/mL after 5 min of exposure to 0.2% Clidox-S at 25°C (Fig. 3A), whereas a lower population reduction of 3.08 log cfu/mL was noted on the control cells of L. monocytogenes exposed to same concentration of Clidox-S (0.2%) for a similar exposure period of 5 min (Fig. 1A). It thus appears that Salmonella Typhimurium is more susceptible than L. monocytogenes to Clidox-S.

Viability of acid-adapted and control Salmonella Typhimurium during exposure to Clidox-S (0.2%) at 25°C
Acid adaptation affects the viability of Salmonella Typhimurium exposed to Quatricide
The change in the viability of Salmonella Typhimurium exposed to Quatricide (0.5%) at 25°C and 40°C is shown in Fig. 4. Regardless of exposure temperature, the viability of the control and the acid-adapted Salmonella Typhimurium decreased with the extension of exposure time. The reduction in viability was more marked with the control cells rather than the acid-adapted cells, and more pronounced at 40°C than at 25°C. Further, it was found that the control cells of Salmonella Typhimurium showed a population reduction of 2.30 log/mL after 50 min of exposure to 0.5% Quatricide at 25°C (Fig. 4A). In contrast, a relatively higher population reduction of 3.37 log cfu/mL was noted with the control cells of L. monocytogenes after same period of exposure to a relatively low dose of Quatricide (0.2%) at 25°C (Fig. 2A). This result further suggested that Salmonella Typhimurium was less susceptible than L. monocytogenes to Quatricide.

Viability of acid-adapted and control Salmonella Typhimurium during exposure to Quatricide (0.5%) at 25°C
The enhanced resistance of acid-adapted cells of test organisms to disinfectants observed in present study is in agreement with the report of Moorman et al. (2005). Moorman et al. (2005) found that exposure of Listeria innocua to acid stress diminished sensitivity to 10 ppm cetrimide, a quaternary ammonium sanitizer, whereas this observed phenomenon is different from the reports of Leyer and Johson (1997) and Stopforth et al. (2003). The former reported that acid adaptation of Salmonella Typyimurium at a pH of 5.0 to 5.8 for one to two cell doublings resulted in marked sensitization of this pathogen to halogen-based sanitizers, including hypochlorous acid and iodine. The latter found that cells of E. coli O157:H7 wash with 2% lactic acid (pH 4.33–4.48) or acetic acid (pH 4.60–4.71) were more sensitive to subsequent sanitizing treatment with 200 ppm QAC. Further, Adrião et al. (2008) reported that acid-adapted cells of L. monocytogenes A9 adherent to polystyrene showed enhanced resistance to hypochlorite, whereas this was not observed with stain T8. Differences in the conditions of acid adaptation and disinfection as well as differences between microbial strain examined may all lead to these discrepancies.
Conclusion
Data collected from the present study demonstrated that acid adaptation enhanced the resistance of both pathogens to the disinfectants examined. L. monocytogenes BCRC 14846 was more susceptible to Quatricide than Salmonella Typhimurium BCRC 10747, whereas the latter was more susceptible to Clidox-S than the former. Further, both disinfectants showed higher antimicrobial activity at 40°C than at 25°C. Thus, more sanitizing effect of Clidox-S and Quatricide can be achieved at 40°C than 25°C. These findings should inform the development of effective disinfection processes that can be used throughout the food supply chain.
Footnotes
Acknowledgment
This research was financially supported by The National Science Council, ROC (Taiwan) (NSC 98-2313-B-002-037-MY3).
Disclosure Statement
No competing financial interests exist.