
Abstract
As part of the C-EnterNet surveillance program of the Public Health Agency of Canada, 122 pooled swine manure samples from 10 farms in Ontario, Canada were collected and tested for Giardia and Cryptosporidium. Giardia duodenalis cysts and Cryptosporidium spp. oocysts were detected using immunofluorescence microscopy. Nested-polymerase chain reaction protocols were performed to amplify the small subunit rRNA gene and the β-giardin gene for G. duodenalis, and the small subunit rRNA gene and the heat shock protein-70 gene for Cryptosporidium spp. The DNA amplicons were sequenced to determine genotypes and species. A mixed multivariable method was used to compare the presence of Giardia and Cryptosporidium in different stages of production. Both Giardia cysts and Cryptosporidium oocysts were present on all tested farms, with 50.8% of the samples positive for G. duodenalis and 44.3% positive for Cryptosporidium spp. by microscopy, and 66.4% and 55.7%, respectively, positive by polymerase chain reaction (PCR). No significant agreement was observed between microscopy and PCR method to detect Giardia and Cryptosporidium (p<0.05). The prevalence of Giardia in manure pits and finisher pigs did not differ (p>0.05), however, it was less frequent (odds ratio, OR=0.21 [0.07, 0.63]) among sows. Cryptosporidium was more likely (OR=3.6 [1.3, 9.9]) to be detected in manure pits and weaners (OR=3.3 [1.1, 10.0]) compared to finisher pigs, and it was less frequent (OR=0.06 [0.007, 0.55]) in sows than in finishers (p<0.05). DNA sequencing demonstrated that 92.1% of the Giardia isolates were Assemblage B and 7.9% were Assemblage E. The most prevalent Cryptosporidium were Cryptosporidium parvum (55.4%), and Cryptosporidium sp. pig genotype II (37.5%). These findings indicate that the occurrence of zoonotic isolates of G. duodenalis and Cryptosporidium is very high on swine farms in southern Ontario, and that there is a potential for transmission between swine and humans by means of cyst and oocyst contaminated water or foods.
Introduction
Numerous studies have demonstrated a high prevalence of both parasites in livestock. Giardia duodenalis and Cryptosporidium spp. have been reported in swine worldwide (Koudela et al., 1991; Villacorta et al., 1991; Quìlez et al., 1996; Atwill et al., 1997; Izumiyama et al., 2001; Wieler et al., 2001; Ryan et al., 2003; Yu and Seo, 2004; Maddox-Hyttel et al., 2006; Vítovec et al., 2006; Chen and Huang, 2007; Hamnes et al., 2007; Zintl et al., 2007; Johnson et al., 2008; Armson et al., 2009; Kváč et al., 2009a; Featherstone et al., 2010; Feng and Xiao, 2011). In Canada, several studies have reported the presence of Giardia and Cryptosporidium in swine (Sanford, 1987; Olson et al., 1997; Guselle et al., 2003; Thatcher and Friendship, 2003; Guselle and Olson, 2005).
Giardia comprises six different species, among which only G. duodenalis has been known to infect multiple host species (Xiao and Fayer, 2008). There has been relatively little molecular characterization done on isolates of G. duodenalis from pigs; however, a few assemblages, particularly E and A have been reported (Ey et al., 1997; Langkjær et al., 2007; Armson et al., 2009). Distribution of the G. duodenalis genotypes may differ geographically and are reported to be age-specific (Ey et al., 1997; Guselle and Olson, 2005; Langkjær et al., 2007). In a study done in Alberta, Canada, pigs were found to be infected with zoonotic G. duodenalis Assemblage A (Guselle and Olson, 2005).
A number of species and genotypes of Cryptosporidium have been reported in pigs (Morgan et al., 1999; Ryan et al., 2003, 2004; Vítovec et al., 2006; Xiao et al., 2006; Hamnes et al., 2007; Langkjær et al., 2007; Zintl et al., 2007; Johnson et al., 2008; Kváč et al., 2009a; Jenkins et al., 2010). Among 18 Cryptosporidium species (Xiao and Fayer, 2008), pigs can be infected with Cryptosporidium suis, Cryptosporidium sp. pig genotype II, and C. parvum (Ryan et al., 2004). In Canada, two studies have identified C. suis in pigs (Guselle et al., 2003; Guselle and Olson, 2005). The Cryptosporidium strains considered to be most infectious to humans are Cryptosporidium hominis, C. parvum, and Cryptosporidium cervine (Leoni et al., 2006). However, C. suis is of concern in terms of zoonotic transmission, as infections have been reported in humans (Xiao et al., 2002; Leoni et al., 2006), as have Cryptosporidium muris infections, which have also been reported in pigs (Gatei et al., 2002; Tiangtip and Jongwutiwes, 2002; Palmer et al., 2003; Zintl et al., 2007; Jenkins et al., 2010).
G. duodenalis and Cryptosporidium spp. infections in swine have generally been reported as being asymptomatic (Sanford, 1987; Koudela et al., 1991; Xiao et al., 1994; Quìlez et al., 1996; Guselle et al., 2003; Ryan et al., 2003; Vítovec et al., 2006; Zintl et al., 2007), whereby apparently healthy pigs may shed infective cysts and oocysts into the environment. Exposure to infective oocysts/cysts through contaminated water and food are important mechanisms of transmission to humans. It is important, therefore, to investigate possible sources of infection in humans. The objective of this study, therefore, was to determine the occurrence, as well as the genotypes and species of G. duodenalis and Cryptosporidium spp. on a subset of Ontario swine farms.
Materials and Methods
Study farms and sample collection
Ten swine farms in Ontario, Canada, were visited three times between September 2005 and May 2006. At each visit, fecal samples were collected from the stored manure pit, and fresh pooled samples were obtained from finisher, sow, and weaner pens. On each farrow-to-finisher farm, one pooled fecal sample was collected from six finisher pens, one pooled sample from six weaner pens, and one pooled sample from six randomly selected sows in the room. In total, 122 pooled fecal samples (31 manure pits, 43 finishers, 24 sows, and 24 weaners) were collected over the entire period of the study.
Sample preparation
The sucrose flotation technique was used as described by Dixon et al. (2011) with the following modifications. Briefly, 20 g of feces were added to 35 mL of phosphate buffered saline with 0.01% Triton X-100 (PBS-TX) pH 7.4, and the slurry was thoroughly mixed with an applicator. The suspension was passed through four layers of gauze and layered over 15 mL of 1 M sucrose solution (specific gravity 1.13) in a conical 50 mL Falcon tube. Since the manure pit sample was liquid, 20 mL was directly layered over the sucrose solution. Samples were centrifuged at 800 g for 5 min. Following centrifugation, the interface and the upper layer of liquid was transferred to a clean tube and re-centrifuged at 800 g for 5 min. The supernatant was decanted and the pellet was re-suspended in 1 mL PBS-TX and used for microscopy and molecular characterization.
Microscopy
For each sample, 100 μL of suspension was transferred to a 1.5 mL microcentrifuge tube. Fluorescein isothiocyanate-labeled monoclonal antibody solutions (25 μL each of Giardi-a-Glo and Crypt-a-Glo; Waterborne Inc., New Orleans, LA) were then added to the tube, which was then incubated at room temperature for 45 min in the dark. After incubation, the excess antibody was washed by adding 1 mL PBS-TX, vortexed and centrifuged at 10,000 g for 10 min. The supernatant was removed and the pellet resuspended in 100 μL PBS. Twenty microliters was placed on a microscope slide and examined as described by Dixon et al. (2011).
DNA extraction
DNA was extracted from each sucrose flotation concentrated sample using the DNeasy Tissue Kit (Qiagen Inc., Mississauga, ON) with a slightly modified protocol as described by Dixon et al. (2011).
Giardia PCR
Nested-PCR was performed to amplify fragments of the small subunit (SS) rRNA gene and the β-giardin gene for Giardia using previously described PCR protocols. Amplification of a 292 bp fragment of the SS rRNA gene was performed as described in Coklin et al. (2007). Amplification of fragments of the β-giardin gene was modified from a method by Cacciò et al. (2002). The PCR mix consisted of 1 H buffer containing 1.5 mM MgCl2, 200 μM of dNTP, 0.5 μM of each primer, 2.5 U of GoTaq polymerase (Promega Corp., Madison, WI) and 10 μL of purified DNA in the first round in a final volume of 50 μL. The PCR cycling consisted of an initial denaturation step of 5 min at 94°C, a set of 35 cycles of 94°C for 30 sec, 30 sec of annealing (58°C in the first round, 66°C in the second round) and 30 sec at 72°C followed by final extension at 72°C for 10 min.
Cryptosporidium PCR
Nested-PCR was performed to amplify fragments of the SS rRNA gene and the heat shock protein-70 gene for Cryptosporidium. Amplification of 825–850 bp fragments of the SS rRNA gene using the primers of Xiao et al. (1999) was performed as described in Coklin et al. (2010). Amplification of 448 and 325 bp fragments of the heat shock protein-70 gene (Guselle et al., 2003) was performed as described in Coklin et al. (2007). All PCR products were electrophoresed on 1.2% agarose gels and visualized with ethidium bromide staining.
DNA sequence analysis
DNA sequencing was performed at the McGill University and Genome Quebec Innovation Centre, Montreal, QC, using a 3730xl DNA Analyser (Applied Biosystems, Foster City, CA). PCR products were purified and sequenced in both directions using the same PCR primers as for the original amplifications. The Genetic Computer Group sequence analysis package (version 10.3, Madison, WI) was used to assemble and analyze the DNA sequences.
Data analysis
Data were entered into a spreadsheet (Microsoft Excel 2003; Microsoft Corp., Redmond, WA) and imported into Stata 11 Intercooled for Windows XP (StataCorp LP, College Station, TX). A mixed multivariable logistic regression modeling method was used to investigate various factors that influence the presence of Giardia and Cryptosporidium on swine farms (Dohoo et al., 2009). In each model, “farm” variable was included as a random effect to take the clustering into account. Also the “stage” variable was included as fixed effect into the models to compare the presence of Giardia and Cryptosporidium among samples collected from manure pits, finishers, sows, and weaners. To control the temporal effect on the presence of the Cryptosporidium oocysts and Giardia cysts, the “visit” variable (visit 1, visit 2, and visit 3) was included into the model if significant at the level of p<0.05. McNemar's χ 2 was used to determine whether there was a difference in the proportion positives of Giardia and Cryptosporidium obtained when testing samples by microscopy compared to PCR.
Results
Both Giardia cysts and Cryptosporidium oocysts could be recovered from at least one of the pooled fecal samples on all 10 farms tested over the three visits of the study. Giardia cysts and Cryptosporidium oocysts were observed by microscopy in 62 (50.8%) and 54 (44.3%) of the pooled samples, respectively. Using PCR, 81 (66.4%) and 68 (55.7%) of the pooled fecal samples were positive for Giardia and Cryptosporidium, in that order. Overall, prevalence of both Giardia cysts and Cryptosporidium oocysts deemed to be higher during visit one (September and October).
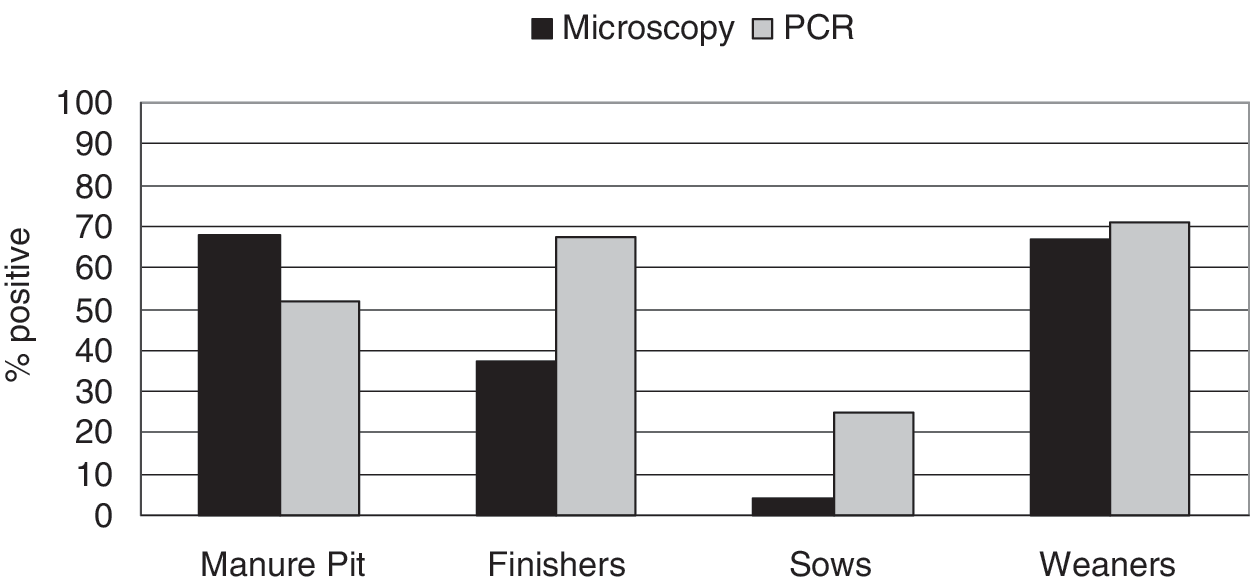
The presence of Giardia and Cryptosporidium detected by microscopy and PCR methods in manure pit, finisher pig, weaner, and sow samples is demonstrated in Figures 1 and 2. The positive proportions of Giardia and Cryptosporidium obtained by microscopy and PCR differed (McNemar's χ 2 p<0.05) indicating that there was no significant agreement between the two testing methods. The presence of Giardia in samples collected from finisher pigs did not differ significantly from that in manure pits or weaners (p>0.05) (Table 1). However, Giardia was detected less frequently from sow samples than from finisher pigs. Giardia oocysts were detected more frequently in the fecal samples collected during visit 1 (September and October) compared to visits 2 and 3 (November to January, and April and May, respectively) (Table 1). Cryptosporidium was detected more frequently from pooled manure pit and weaner samples than from finisher pig samples (Table 1). However, it was detected less frequently from samples from sows compared to finisher pig samples. The time of visit was not significantly associated with prevalence of Cryptosporidium oocysts.

Presence of Giardia detected by microscopy and polymerase chain reaction (PCR) in fecal samples.
Presence of Cryptosporidium detected by microscopy and PCR in fecal samples.
Visit 1 (September to October, 2005), visit 2 (November 2005 to January 2006), visit 3 (April to May 2006).
“Visit” variable was not associated with presence of Cryptosporidium oocysts in fecal samples and therefore was not included in the final model.
CI, confidence interval.
Of the 81 samples positive for G. duodenalis by PCR, DNA sequences were obtained for 63, including 16 from manure pits, 24 finishers, 10 sows, and 13 weaners. For Cryptosporidium spp., sequence data were available for 56 isolates recovered from 54 samples, including 12 manure pits, 23 finishers, 5 sows, and 14 weaners. Giardia and Cryptosporidium genotypes are shown in Tables 2 and 3.
DNA sequences were available for 63 isolates recovered from 16 manure pits, 24 finishers, 10 sows, and 13 weaners.
DNA sequences were available for 56 isolates recovered from 54 samples including 12 manure pits, 23 finishers, 5 sows, and 14 weaners.
Discussion
A large proportion of the pooled swine manure samples tested in the present study showed the presence of G. duodenalis cysts and Cryptosporidium spp. oocysts. Generally lower prevalences for both parasites have been previously reported in swine worldwide (Johnson et al., 2008; Armson et al., 2009). Further, a greater proportion of the samples from the younger animals were positive for both G. duodenalis and Cryptosporidium spp. in the present study than those samples collected from the sows. This finding was in agreement with previous studies which have reported the prevalence in swine to be age-specific, with a higher prevalence being reported in young animals (Xiao et al., 1994; Quìlez et al., 1996; Olson et al., 1997; Maddox-Hyttel et al., 2006; Zintl et al., 2007), but is in contrast with the findings of an earlier Canadian study (Olson et al., 1997). It is possible that infected sows may shed a lower number of oocysts and cysts compared to finishers and weaners, resulting in fewer positives being detected by microscopy methods. This would be consistent with the observation that PCR detected more positive samples compared to microscopy (Ryan et al., 2005).
In the present study, G. duodenalis Assemblage B predominated in all swine age groups, and to our knowledge, Assemblage B has only previously been reported in a single pig manure sample in Europe (Feng and Xiao, 2011). Since both G. duodenalis Assemblages A and B are commonly reported in humans, the predominance of Assemblage B in the present study suggests that swine may have a greater zoonotic potential for G. duodenalis than previously thought.
Similar to the present study, Featherstone et al. (2010) reported the pig genotype II and C. parvum as the most common Cryptosporidium isolates in swine in East Anglia, UK. C. parvum, which is generally recognized as the major zoonotic species, and has been rarely reported in pigs (Kváč et al., 2009a), was common to all age categories, suggesting a zoonotic risk from swine. This finding was in contrast to that of Johnson et al., (2008) who did not detect any C. parvum in pigs in Western Australia and concluded that domestic pigs do not pose a significant public health risk. These authors reported only the presence of C. suis and pig genotype II. The second most frequent Cryptosporidium isolate found in the present study was Cryptosporidium sp. pig genotype II, which is generally considered to be a nonzoonotic genotype, although a recent study by Kváč et al. (2009b) reported this genotype in an immunocompetent man in the Czech Republic. Similarly, C. muris, which was recovered only from manure pits in this study has been reported in human cryptosporidiosis cases (Gatei et al., 2002; Tiangtip and Jongwutiwes, 2002; Palmer et al., 2003). The presence of C. muris in pig manure samples has been previously reported by Jenkins et al. (2010) and Zintl et al. (2007). It suggests the presence of rodents on the farm, and the possible transmission of Cryptosporidium spp. between rodents and pigs, which has been previously suggested by Chen and Huang (2007). C. suis may also be considered to be a zoonotic species as it has been reported in a small number of human cases (Xiao et al., 2002; Leoni et al., 2006). Studies done in Canada and the Czech Republic reported C. suis as the only species present in swine operations (Guselle and Olson, 2005; Vítovec et al., 2006). However, this species was only rarely identified in swine manure samples in the present study.
It is not clear whether the zoonotic G. duodenalis Assemblage B and C. parvum are naturally circulating among swine in this region or whether there may be other external sources of infection. For example, the predominance of Assemblage B in swine in the present study is also suggestive of zooanthroponotic transmission, potentially by means of sewage contaminated water used to water animals and clean facilities.
In the present study, G. duodenalis and Cryptosporidium spp. were detected in a large proportion of the pit manure samples tested. Neither G. duodenalis cysts nor Cryptosporidium spp. oocysts in swine manure are environmental concerns if the manure is handled responsibly (Olson and Guselle, 2000). However, manure management procedures may not be effective in eliminating zoonotic agents from hog manure, and as such they may fail to prevent the environmental contamination (Jenkins et al., 2010; Ziemer et al., 2010). Létourneau et al. (2010) have found zoonotic pathogens in the manure of pig operations following various manure management practices. Thurston-Enriquez et al. (2005) have reported high run-off concentrations of Cryptosporidium spp. oocysts and G. duodenalis cysts from experimental plots treated with swine slurry, and suggested that infective Cryptosporidium oocysts and Giardia cysts may be transferred to ground water when precipitation creates run-off from manure on agricultural fields. Similarly, Langkjær et al. (2007) concluded that untreated pig manure used on open fields could be a concern in the environmental transmission of potentially zoonotic Cryptosporidium species. A recent spatial epidemiological study has shown that C. parvum infections in humans were more frequent in areas where private water supplies are more common, and the ratio of farm density to human density is higher (Pollock et al., 2010). Both G. duodenalis and Cryptosporidium spp. are highly resistant to chlorination, and contaminated water is a major contributor to human infections. Therefore, it is important to consider manure management systems and disinfection methods in the effective inactivation of parasites in hog manure.
G. duodenalis and Cryptosporidium spp. infections in swine have generally been reported as being asymptomatic (Sanford, 1987; Koudela et al., 1991; Xiao et al., 1994; Quìlez et al., 1996; Guselle et al., 2003; Ryan et al., 2003; Vítovec et al., 2006; Zintl et al., 2007), although Cryptosporidium may be a copathogen in diarrheal disease in neonatal and weaning pigs (de Graaf et al., 1999). Additionally, subclinically infected pigs may experience slower weight gain (Bilic and Bilkei, 2006). Therefore, in addition to public health concern, the presence of Cryptosporidium and Giardia may be clinically and economically important in pigs (Thatcher and Friendship, 2003) and there may be an economic incentive for producers to control these parasites.
The occurrence of both G. duodenalis and Cryptosporidium spp. was high in the swine manure samples in the present study, and zoonotic genotypes and species were identified. Further study will be required to identify the source of these infections in swine, and molecular characterization of human isolates in this region would be required to investigate the actual risk of transmission to humans.
Footnotes
Acknowledgments
We would like to acknowledge the Public Health Agency of Canada for funding the C-EnterNet program. We thank Bryan Bloomfield at the University of Guelph, Department of Population Medicine for the field work, as well as the pork producers for their participation in this study.
Disclosure Statement
No competing financial interests exist.