
Abstract
Salmonellosis is one of the major concerns in the poultry industry and some serovars of Salmonella involve in zoonosis. This study determines the seroprevalence of Salmonella in poultry and their drug-resistant patterns, variability in infectivity and mortality rate of birds, and predilection of some serovars to cause zoonoses. The average seroprevalance of Salmonella in three different age groups was found to be 37.9%. A total of 503 samples were examined over a period of 1 year from five different poultry farms of a semiurban area of Savar, Dhaka, Bangladesh. The prevalence of Salmonella was recorded to be 21.1%. Salmonella was found high in dead birds (31.2%) than live birds (18.1%). Salmonella infection was higher (23.6%) in summer than in winter (12.9%) season. Among the 106 isolates, 46 belong to serogroup B (43%) and 60 isolates to serogroup D (57%). The highest Salmonella infection was recorded as 47.9% on the 30–35-week-old birds. A total of 106 Salmonella isolates were used for antimicrobial susceptibility test against 10 common antibiotics and 17 multiple drug resistance patterns were found. Among the isolates, 69 (65%) harbored plasmids 1–4 with size variation between >1.63 and >40 kb and rest 37 (35%) isolates were plasmid free but showed resistance against 5–10 antibiotics. The results of the present investigation suggested that multiple drug resistance is common among the Salmonella isolates of poultry and some of these isolates may have zoonotic implications.
Introduction
E
Poultry is known to be the largest single reservoir of Salmonella (Gupta et al., 1999). Salmonellosis is one of the most important diseases in poultry that cause serious economic loss because of mortality and reduced egg production (Khan et al., 1998). The major host-adapted serovars of Salmonella in poultry are Salmonella Gallinarum and Salmonella Pullorum, which are responsible for fowl typhoid and Pullorum disease, respectively (Snoeyenbos, 1994; Khan et al., 1998). In addition to avian salmonellosis (Palaniswamy et al., 1989; Verma and Gupta, 1997), Salmonella Typhimurium and Salmonella Enteritidis are unambiguously categorized as zoonotic hazards, because poultry are known to be the major transmitter of nonhost-adapted salmonellosis in humans (Rahman et al., 1997).
Salmonella infection is one of the major constraints of poultry farming that hindered its development in Bangladesh (Kamaruddin and Giasuddin, 2003; Das et al., 2005). In recent days, the prevalence of salmonellosis in breeder flock, commercial broiler, and layer flocks is increasing day by day. However, very limited research works had been carried out in Bangladesh concerning salmonellosis in poultry so far. Therefore, salmonellosis status of a farm needs to be determined for its proper control and management (Ahmed et al., 2008). The objective of this study is (1) to determine the seroprevalence of Salmonella in poultry, (2) antibiogram of isolated salmonella strains, (3) variability in infectivity and mortality rate in differing season, and (4) predilection of some serovars to cause zoonoses.
Materials and Methods
Study area
This study was conducted in five different layer poultry farms located in semiurban area of Savar region of Dhaka, Bangladesh (Fig. 1), during the period May 2009 to June 2010. The samples were collected from the birds of selected poultry farm and brought to the Department of Microbiology, University of Dhaka, for laboratory analysis.
Sample collection location of five poultry farms in Savar area indicated as A, B, C, D, and E.
Seroprevalence study
Sample collection
A total of 346 blood samples of the wing vein of individual birds of 20th–25th, 30th–35th, and >40th weeks of age were collected from four selected poultry farm (10% of the total flock). Blood samples were aseptically collected in sterile vial with sterile 5-mL syringe and the samples were allowed to clot in the syringe and kept for 1–2 hours at room temperature. After clotting, sera were separated, centrifuged (12,000 rpm, 1 minute) at room temperature and poured in sterile vials, individually labeled, and stored at −20°C until further use.
Salmonella antigen
Standard Salmonella Pullorum antigen manufactured by Lohman Animal Health Ltd. was used for serum plate agglutination (SPA) test for the detection of Salmonella antibodies in the sera samples.
Detection of Salmonella infection by SPA test
The SPA test was performed according to the procedure described in OIE Manual, 2000, with crystal violet-stained Salmonella antigen (Pullorum antigen; Lohman Animal Health Ltd.). For this test, 0.02 mL of antigen and 0.02 mL of chicken sera were placed side by side with a micropipette on a glass plate. Then, the antigen and the sera were thoroughly mixed by stirring with a small tooth pick followed by rocking. Results of SPA test were read within 2 minutes. In positive case, granules were slowly formed, indicating that sera samples contained antibody against Salmonella infection. In negative case, granules were not formed within 2 minutes, indicating that antibody against Salmonella were absent in the sera samples. The results of SPA test were recorded.
Isolation and identification of Salmonella
Sample collection
A total of 391 cloacal swabs were collected from live birds and a total of 112 samples were collected from liver, intestine, ovary, oviduct, and spleen swab after postmortem examination of dead birds. Sterile cotton swab sticks were used for sample collection and collected samples were directly brought to the laboratory in an insulated ice box with minimum delay and bacteriologically examined immediately. Isolation and identification of salmonellae were done according to the procedure described by OIE (2000), Merchant and Packer (1967), and Cowan (1985).
Cultivation of the sample
The collected swab containing samples were grown into tetrathionate broth (Oxoid Ltd.) at 37°C for 18–24 hours. Tetrathionate broth-grown cultures were grown in MacConkey, brilliant green, and Salmonella–Shigella agar to get pure and putative Salmonella culture. The organisms were further characterized as Salmonella species according to their morphology, Gram staining, mortality, and biochemical properties (Merchant and Packer, 1967; Cowan, 1985; OIE, 2000).
Serogrouping of Salmonella isolates
Serogrouping of Salmonella isolates was performed by slide agglutination test using commercial Salmonella-specific polyvalent O (A-I) antisera, Salmonella O group B (Factor O: 4, 5, 27) antisera, and Salmonella O group D (Factor O: 9, 46) antisera (S&A Reagent Lab). The test was performed according to the protocol supplied by the manufacturer.
Antibiogram study of Salmonella isolates
In vitro antibiotic sensitivity test of the isolated Salmonella was performed through disc diffusion method (Bauer et al., 1966) with the standard commercial discs manufactured by Oxoid Ltd. In this method, Salmonella isolates were grown overnight on brilliant green agar (Oxoid Ltd.) at 37°C and the overnight grown isolates were then inoculated into nutrient broth and incubated at 37°C for 18–24 hours. After incubation, one loopful of inocula was poured onto 9 mL of Mueller Hinton broth (Oxoid Ltd.) and incubated at 37°C for 5–6 hours. The bacterial cultures were compared with McFarland (Jorgensen et al., 1999) turbidity standard (108 CFU/mL) and streaked evenly in three planes onto the surface of the Mueller Hinton agar plate (5×40) with a sterile cotton swab. After the inocula dried, antibacterial discs such as ampicillin (A) 10 μg, amoxycillin (Am) 10 μg, ciprofloxacin (C) 5 μg, erythromycin (E) 15 μg, gentamycin (G) 10 μg, penicillin-G (P) 10 units, streptomycin (S) 10 μg, sulfamethoxazole (Su) 25 μg, tetracycline (T) 30 μg, and nitrofurantoin (F) 300 μg were placed on the agar aseptically and kept at 4°C for 30–60 minutes for better diffusion. The inoculated plates containing the discs were incubated in an upright position at 37°C overnight and/or 24–48 hours (if necessary). The results were expressed as the diameter of inhibition zone around the paper disk (8 mm).
Maintenance of stock culture
The stock cultures of the Salmonella isolates were preserved in TSB medium containing 20% glycerol and kept at −86°C. Working cultures were kept at 4°C on tryptic soy agar slants (Nissui) and were periodically transferred at 15 days interval to fresh slants.
Isolation of plasmid DNA
A single bacterial colony was transferred into screw-capped tubes containing Luria broth medium 1:5 (v/v) containing appropriate antibiotics and incubated at 37°C overnight with shaking. The culture was pelleted by centrifugation (12,000 rpm, 30 seconds) at 4°C. The supernatant was removed and the pellet was homogeneously suspended in buffer and plasmids were isolated according to Birnboim and Doly (1979) method or using PureLink™ Quick Plasmid Miniprep kit (Invitrogen). A 5 μL plasmid DNA was loaded on to a 1.0% agarose gel containing 0.5 μg mL−1 ethidium bromide and electrophoresed in 1×Tris–boric acid–EDTA buffer. The plasmid DNA were visualized by an ultraviolet transilluminator and recorded using the Alfa Digital documentation imaging system (Alfa Innotech Corporation).
Statistical analysis
The serological test results were statistically analyzed based on farm location and age and sex of the birds. Data were subjected to chi-square statistics using Microsoft Excel program. Significant differences of the data were established by least significant difference at the 5% level of significance.
Results and Discussion
The SPA test revealed that the average seroprevalance of Salmonella in three different age groups was 37.9% (Table 1). The average seroprevalence was lowest between 20 and 25 weeks and recorded as 27.7%, whereas the highest reached 43.8% of 30–35-week-old birds. No significant difference of the prevalence among five farms and three groups of ages were observed in chi-square test. Seroprevalence study under various conditions of varying season and age and sex of birds have been carried out by different researchers. The overall prevalence of Salmonella was reported to be as low as 14.1% to as high as 45.9% (Islam et al., 2006; Ahmed et al., 2008; Hossain et al., 2010). In another study, the mean seropositivity of different farms in three different age groups was found to be 18.9%±2.3%, 33.2%±3.53%, and 27.8%±2.67% on 10th, 24th, and 40th week of age (Hossain and Islam, 2004). Similar results by other workers also found that the prevalence of Salmonella infection increases with increasing age (Sikder et al., 2005; Islam et al., 2006).
A total of 503 poultry samples were analyzed; of them, 416 were live poultry and 87 were dead poultry. Of 416 live poultry samples, 71 (18%) were found to be Salmonella positive, whereas from 87 dead poultry, 35 (31%) poultry was Salmonella positive (Table 2). Significant difference of the prevalence between live poultry and dead poultry (p<0.01) was observed by chi-square test. It should be noted that the live poultry purchased for this study was not clinically diseased; on the other hand, the dead poultry purchased for this study were case fatalities. The isolation rate of Salmonella had been reported to be 24.5%, 28%, 47.6%, and 14.3%, respectively, from the liver of dead poultry (Rusul and Yassin, 1996; Habib-ur-Rahman et al., 2003; Lee et al., 2003; Tibaijuka et al., 2003). These differences in isolation rate may be due to the geographical variation.
The prevalence of Salmonella isolates within a 1-year period has been presented in Table 3. The average prevalence of Salmonella infection was found higher (23.6%) in summer season than in rainy (21.3%) and winter (12.9%) seasons (Table 3). These results are in agreement with the result reported by Saleque et al. (2003), Rahman et al. (2004), and Sikder et al. (2005). Recently, Hossain et al. (2010) reported that the prevalence of Salmonella infection was the highest (18.5%±11.9%=30.4%) in summer, followed by winter (11.6%±12.1%=23.7%), rainy season (14.2%±10.8%=25.0%), and autumn (13.3%±10.0%=23.3%).
From the 503 poultry samples, 106 Salmonella were isolated (Table 4). Among the 106 isolates, 46 belonged to serogroup B (43%) and 60 isolates to serogroup D (57%) (Table 5). The most prevalent serogroup identified in this study was serogroup D. These findings are in agreement with the result reported by Arroyo and Arroyo (1995).
P, penicillin 10 units; Amp, ampicillin 10 μg; A, amoxicillin 25 μg; E, erythromycin 15 μg; S, streptomycin 10 μg; T, tetracycline 10 μg; F, nitrofurantoin 300 μg; Su, sulfamethaxol 10 μg; G, gentamycin 10 μg; C, ciprofloxacin 5 μg. Antibiotic per disc.
Salmonella isolates that showed at least one variable (farm, serogroup, and plasmid profile) different are listed in table.
Antibiogram of the 106 isolates against 10 commonly used antibiotics revealed 17 different phenotypic expressions acquiring 10 to 5 drugs resistance concurrently. The prevalence of the multiple drug resistance (MDR) (resistance to greater than or equal to three classes of antimicrobial agents) Salmonella isolates against 10 to 5 antibiotics were 26.4%, 17%, 20.7%, 2.8%, 27.4%, and 5.7% (Table 4). Interestingly, same serogroup with identical MDR phenotypes and plasmid profiles containing Salmonella isolates were found in five farms, indicating the possibility of intrafarm transmission of this bacterium. The relative susceptibility of commonly used antibiotics revealed that the highest percentage of antibiotic resistance was found against penicillin-G and ampicillin, amoxicillin, tetracycline, nitrofurantoin, sulfamethaxol, gentamycin, and ciprofloxacin, with 100%, 99%, 98%, 93%, 78%, 60%, 46%, and 40%, respectively (data not shown). Similar resistance patterns to common antibiotics and concurrent increase in multiple resistances in Salmonella isolates worldwide had been reported elsewhere (Verma et al., 1993; Anjanappa et al., 1994; Hui and Das, 2001) for the last three antibiotics (Molla et al., 2003; Carraminana et al., 2004; Muhammad et al., 2009; Maripandi and Al-Salamah, 2010). The multiple antibiotics resistances observed in the present study is an alarming sign for public health concern, because these antibiotics are frequently used for the treatment of human infection diseases. Increasing resistance to important antimicrobials used for human therapy, such as cephalosporins and fluoroquinolones, as well as increasing multiple resistance often linked with virulence determinants is an increasing concern (Su et al., 2008; Garcia-Fernandez et al., 2009). The high MDR index (1–0.5) found for 10 commonly used antibiotics (Table 4) indicated the alarming situation and implicated that the Salmonella isolated sites were highly contaminated with large numbers of these antibiotics.
To verify the MDR properties whether chromosomal or plasmid borne, the plasmid profiles of 106 Salmonella isolates were examined and the results are shown in Table 5 and Figures 2 and 3. Plasmid profile analysis revealed that 69 isolates among the total 106 contained 1 to 4 plasmids of varying sizes (>1.63 to >40 kb) (Table 5); but rest of the 37 isolates were plasmid free and showed resistance against 5–10 antibiotics with 12 different resistance patterns (Tables 4 and 5). Plasmid profiles of 46 isolates belonging to serogroup B revealed that 76% (35 of 46) of the isolates contained plasmid, whereas this value was 56.67% (34 of 60) for serogroup D (Figs. 2 and 3). Surprisingly, 34 isolates belong to both B and D serogroups, contained a single large plasmid of >40 kb, and showed six resistance patterns. Isolate number 90 and 94 harbored the large plasmid, >40 kb, along with three other small plasmids of 4, 3, and 2 kb, although these two isolates show different antibiograms and belong to different serogroups. In contrast, isolate number 72 belonging to same serogroup and with similar MDR pattern with the isolate number 90 harbored different plasmid profiles (>40, 3, and 2 kb). On the other hand, Salmonella isolated from same farm, belonging to same serogroup, and having similar plasmid profile also showed different antibiograms. Salmonella isolated from same or different farm belong to same or different serogroup and had not showed same plasmid or different antibiograms. In contrast, it was observed that Salmonella isolated from same farm that belonged to the same serogroup and showed same antibiograms varied in plasmid profiles or even did not contain any plasmid. The same was also true for cross-serogroups.
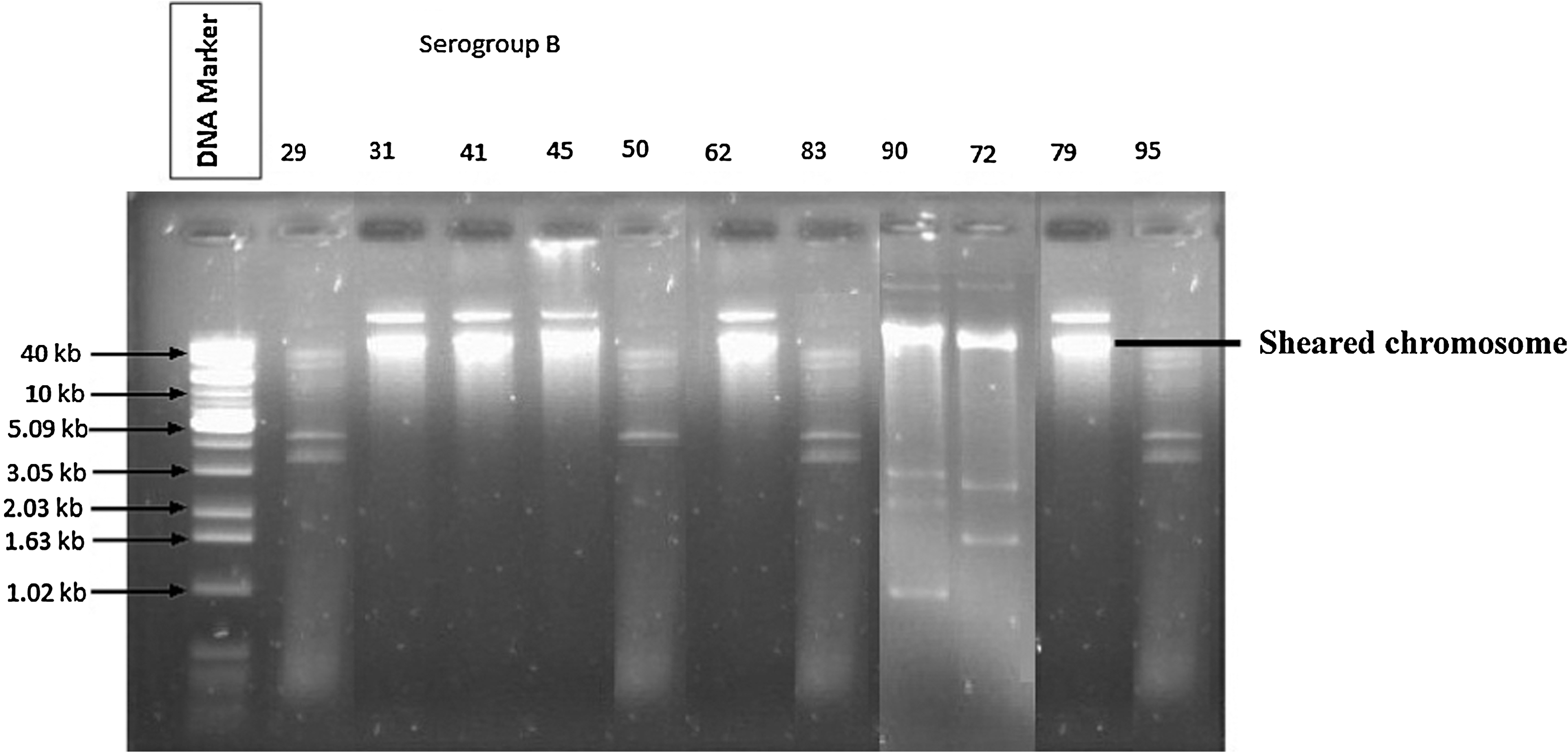
Plasmid profile of representative 11 Salmonella isolates of serogroup B. Plasmid pattern of 11 isolates (29, 31, 41, 45, 50, 62, 83, 90, 72, 79, 95) of Salmonella are shown in lanes 2–12, and lane 1 is the marker used.

> Plasmid profile of representative 11 isolates of serogroup D. Plasmid pattern of 11 isolates (14, 20, 26, 33, 39, 67, 89, 93, 94, 97, 98) of Salmonella are shown in lanes 2–12, and lane 1 is the marker used.
The present work was designed to investigate salmonellae involved in poultry infection on the basis of five variables farm code (situated within 5 sq. miles), serotypes, resistotypes, and plasmid profiles. Depending on at least single variable variation, 56 Salmonella isolates were identified with 17 different resistotypes harboring 1–4 plasmid or none (Table 5). The very high unanticipated levels of commonly used therapeutic antibiotics resistance observed in Salmonella isolates is probably due to the indiscriminate and widespread use of these antibiotics in the poultry, veterinary, and public health practices, because people have very easy access to various antimicrobials in Bangladesh.
Salmonella harbored multiple R-plasmids as reported by Maripandi and Al-Salamah (2010), Morshed and Peighambari (2010), Singh et al. (2010), and Threlfall et al. (2005). The study indicated that the presence or number of plasmids is not associated with resistant phenotype of Salmonella isolates (Table 5). The absence of correlation between MDR patterns and the plasmid profiles in Salmonella isolates indicates that the origin of the resistance properties might be on chromosome. Similar results of Salmonella isolates from veterinary origin in India were reported by Singh et al. (2010). Alarmingly, 35% of the Salmonella isolates were plasmid free, but had resistance to 5–10 antibiotics and belonged to 3–7 different antimicrobial groups. So far, to the best of our knowledge, this is the first study reporting Salmonella resistant to 10 antibiotics and belonging to 7 antimicrobial groups. Salmonella Tyhimurium DT104 resistance to five antibiotics and belonging to four antimicrobial groups had been documented (Threlfall et al., 2005). Very recently, a plasmid-free Salmonella Anatum strain had been isolated from buffalo sources in India, which is resistant to seven antibiotics and belongs to four antimicrobial groups (Singh et al., 2010). Chromosome-borne bacterial resistance mostly occurs because of alteration of antimicrobial drug target and alteration and/or limit permeation/efflux of the drugs. The alteration of target and drug efflux mechanism had been documented for Salmonella (Threlfall et al., 2005; Nair et al., 2006).
In conclusion, Salmonella isolates from poultry farm of Savar area, Dhaka, Bangladesh, showed MDR properties at alarming levels. Further, a wide range of MDR has implications for veterinary and human therapy, as their misuse in poultry could lead to emergence of resistant zoonotic pathogens. Therefore, MDR strains of Salmonella of poultry origin might be important for public and personal health, as well as for epidemiologists monitoring the spread of MDR in zooonotic Salmonella pathogens in Bangladesh and beyond the borders.
Footnotes
Acknowledgments
The authors appreciate the financial supports for this work provided by Ministry of Education and Ministry of Science and Information and Communication Technology, Government of the People's Republic of Bangladesh. The author also thanks Banglabandhu scholarship authority for granting financial support. The authors thank Salma Akter and Esfat M. Saim Bristy for providing assistance during experiment.
Disclosure Statement
No competing financial interests exist.