
Abstract
Dried grapes (raisin) may carry a significant mycological load contaminated via cultivation, postharvest processing practices, and drying processing. The contamination of raisin with fungi can accelerate spoilage or illness, if pathogens are present. Since raisins are used as food additives in many dishes in Saudi kitchen, there are healthy concerns on the safety of raisins consumed. In this article, the mycological profile of raisins sold in different markets at Jeddah, Kingdom of Saudi Arabia, was studied. The black raisin samples showed high fungal load compared with the white samples. Aspergillus, Alternaria, Cladosporium, Epicoccum Fusarium, and Rhizopus were the most prevalent genera isolated from raisin samples. Among six Aspergillus species isolated in this study, As. carbonarius and As. niger were the most frequently isolated species. Ochratoxin A (OTA) production in raisins was investigated using a combination of chromatographical (thin layer) and molecular (random amplified polymorphic DNA and Multiplex polymerase chain reaction) techniques. The OTA was detected in 70% of the raisin samples. Also, As. carbonarius (14 out of 19 isolates) and As. niger (2 isolates out of 9) were recognized as potential producers for OTA. Also, some molecular markers for detecting the contamination of raisin samples with OTA directly without isolating the producers were tested.
Introduction
Mycobiota and ochratoxin of dried grapes
Molecular detection of ochratoxin
The traditional schemes for the isolation and identification of ochratoxigenic fungi from food samples are time consuming and require a high knowledge of fungal taxonomy. Even with taxonomic expertise, identification is commonly difficult in some genera of fungi that contain a large number of closely related species. Aspergillus Section Nigri includes several species difficult to be identified with traditional methods, because differences are mainly based on the uniseriate and biseriate condition of the sterigmata of conidial heads and on the size and roughness of the conidia (Battilani et al., 2008). Hence, it is imperative to develop methodologies that are relatively rapid, highly specific, and as an alternative to the existing methods. The application of molecular biology techniques can help overcome these problems, because it can reduce the time for identification from days to hours and also allow precise species identification (Sartori et al., 2010). Polymerase chain reaction (PCR)-based methods that target DNA are considered a good alternative for rapid diagnosis because of their high specificity and sensitivity (Accensi et al., 1999; Perrone et al., 2004), especially when multi-copy sequences are used to develop species specific primers (Bluhm et al., 2002). Patino et al. (2005) developed two PCR assays to detect As. carbonarius and As. ochraceus, considered the main sources of OTA contaminating commodities, particularly grapes, coffee, and derivatives, in warm climates. Random amplified polymorphic DNA (RAPD) or amplified fragment length polymorphism have been successfully applied for revealing specific marker sequences, such sequences have been used to design species-specific primers that allow the identification and detection of some ochratoxigenic species in food samples (Fungaro et al., 2004; Niessen et al., 2005; Niessen, 2006; Sartori et al., 2006).
Saudi Arabia is one of the most important raisin markets in the world with an annual consumption of many tones. However, the mycological studies inspecting the fungal contaminants and safety of the related products are limited. Therefore, the aims of this work were to determine the occurrence and load of fungi, the important food borne pathogens, in particular, in raisins offered for sale to consumers in the retail stores at Jeddah region in Saudi Arabia; to specifically analyze the occurrence of Black aspergilli group in these samples; to investigate the occurrence of OTA using chromatographically and molecularly techniques; and to directly test more reliable methods for detecting OTA in the raisin samples.
Materials and Methods
Mycobiota determination
Fifty raisin samples of different origin were collected from different markets in Jeddah City. The samples were surface sterilized and placed on malt extract and dichloran-rose bengal chloramphenicol (DRBC) agar medium (King et al., 1979). The plates were incubated at 25°C for 7 days, and fungal colonies growing on these plates were purified and identified by classical taxonomic methods. From media plates, only colonies belonging to black Aspergilli were transferred to slants to ensure precise counting and for identification to species level (Klich and Pitt, 1988). These isolates were preserved at −80°C for later studies.
Determination of ochratoxin production (thin-layer chromatography)
For the determination of ochratoxin production by black aspergilla strains, the method of Davis et al. (1969) was used.
Detection of natural occurrence of OTA in raisin samples
Raisin samples that showed high contamination with black aspergilla went for OTA detection. Each sample was mixed, homogenized, placed in a glass bottle, and stored at 2–8°C until further analysis. For the qualitative analysis of ochratoxin, thin-layer chromatography (TLC) technique was used. The OTA was detected in the sample extracts after TLC on 0.3 mm layers of Adsorbosil 5 silica gel, using tolueneethyl acetate-90% formic acid (6: 3: 1 v/v/v) for development. Each extract formed a green fluorescent spot under ultraviolet light with the same R r (0.55) as authentic OTA. The fluorescence turned blue on treatment with ammonia (Shotwell et al., 1969).
DNA isolation from fungal isolates
The isolation of DNA from mycelia of As. carbonarius and As. niger was performed according to the method described by Accensi et al. (2001).
DNA extraction from fungal-contaminated raisins
The contaminated raisins were immersed in a 0.4% hypochlorite solution for 2 min. Then, they were rinsed with sterilized distilled water for 1 min. This washing procedure was repeated thrice. To extract the DNA, a CTAB protocol (Doyle and Doyle, 1987) was used. The raisins were first freeze dried and ground using a blender to a fine powder macerated with the addition of liquid nitrogen and protocol of Sartori et al. (2006) was used.
Multiplex PCR reaction using artificially contaminated raisins
The total DNA obtained from one raisin was serially diluted to 101, 102, and 103 in distilled water and used as a template DNA in the multiplex PCR reaction. The PCR reaction mixture contained 3.0 μL DNA of each template, 0.5 μL MgCl2 (50 mM), 2.5 μL Taq DNA polymerase buffer, 1.0×, 1.0 μL primer (10 pmol/μL each), 10.9 μL H2O, 0.1 μL Taq polymerase (5 U/μL), and 2.0 μL dNTP (2.5 mM). Details of cycling conditions and primers were shown in the work of Sartori et al. (2006).
Multiplex PCR assays in naturally contaminated raisins
Raisins obtained from markets were immersed in a 0.4% hypochlorite solution for 2 min and rinsed with sterilized distilled water for 1 min. The samples were put onto soaked filter paper disks inside Petri dishes and incubated at 28°C for 24 h. DNA extraction was performed as just described.
Genetic diversity of As. carbonarius isolates
For molecular analysis, 10-mers were used as random primers in the RAPD procedure. Amplifications were carried out as described by Fungaro et al. (1996).
Computer-assisted analysis with the TREECON for Windows v.1.3b
The results went through the computer-assisted analysis, and the produced matrices will be analyzed by TREECON for Windows (version 1.3b, 1998) wherein the evolutionary tree dendrogram is constructed and produced (Van de Peer and Wachter, 1994).
Results and Discussion
Mycobiota and potential OTA producing fungi in raisin samples
Table 1 showed the mean values of total fungal counts and the Aspergillus spp. counts in malt extract agar (MEA) and DRBC culture media using the direct plating technique. Mean total fungal counts and Aspergillus spp. counts obtained with DRBC culture medium were higher than the mean values obtained with the MEA culture medium. This results may be attributed to the presence of dichloran in DRBC culture medium, which suppressed zygomycetes fungi and, thus, give the chance for other fungi to appear. Varga et al. (2006) reported that seven samples were heavily contaminated by zygomycetes fungi which overgrew other filamentous fungi on MEA plates. The dichloran-rose Bengal medium (King et al., 1979) was successfully used to identify other contaminants in even these samples.
CFU, colony forming units; MEA, malt extract agar; DRBC, dichloran-rose bengal chloramphenicol.
The black fruit samples showed high contamination in comparison with white fruit samples. Mean values of total fungal and Aspergillus spp. counts from black and white fruit were 658.5 and 519; and 525.3 and 390.8 CFU/g, respectively (Table 1). This result was in agreement with results reported by Magnoli et al. (2004) during their work in dried vine fruit from Argentina. Recently, Palumbo et al. (2011) isolated 400 strains from Aspergillus from forty raisin clusters sampled from four California vineyards in the San Joaquin Valley.
The results of Table 2 showed predominant mycobiota of raisin samples, taking into account their occurrences and abundance, respectively, and belonged to Aspergillus spp: (72% and 45.73%) Alternaria alternata (52% and 11.40%), Cladosporium cladosporioides (48% and 25.35%), Epicoccum purpurascens (36% and 1.69%), Fusarium oxysporum (36% and 1.74), Rhizopus stolonifer (36% and 0.87%) and, in a lesser extent, Dreschlera spicifera (26% and 2.12), Penicillium spp. (24% and 6.52%), Humicola gresia (20% and 0.72%), Trichoderma harzianum (16% and 0.48%), and Ulocladium atrum (12% and 0.92%). Alternaria, Aspergillus, Cladosporium, and Penicillium species have been reported as the predominant mycobiota in harvested grapes from Brazil (Da et al., 2002), from France (Sage et al. 2002), and also during ripening of grapes from Spain (Bau et al., 2005). Serra et al. (2005) reported that the most frequent genera isolated from grapes from Portugal were Cladosporium (25%), Alternaria (24%), Botrytis (15%), Penicillium (9%), and Aspergillus (8%).
The most frequent isolated species were As. carbonarius and As. niger. The occurrences and abundance of As. carbonarius and As. niger were (38% and 31.4%) and (18% and 11.59%), respectively (Table 2). The high rate of appearance of As. carbonarius in hot and dry regions is attributed to the resistance of the black Aspergilli to sunlight and ultraviolet light (Benford et al., 2001). Previous mycological studies have reported black aspergilli as the predominant fungi with occurrences between 33 and 100% in raisins (Abarca et al., 2003, Magnoli et al. 2004); and within them, As. carbonarius and As. niger have been recorded between 69 and 100% of vineyards in a study carried out in Australia (Leong et al., 2004). Magnoli et al. (2004) studied 50 samples of dried vine fruit (31 black,19 white) from markets in Mendoza and San Juan Provinces, Argentina. Their results showed that Aspergillus spp. were predominant. The frequency of isolation from black and white dried vine fruits samples were 95% and 41% on DRBCand 75% and 33% on DG18 (Dichloran 18% Glycerol medium), respectively. Also, they reported that the predominant species isolated were As. niger var. niger (75% and 65%), As. niger var. awamori (80% and 55%), and A. carbonarious (45% and 15%) from black dried vine fruits samples in DRBC and DG18, respectively. These species were isolated in lower frequency from white dried vine fruit. In a survey of six vineyards in Australia, soil and vine remnants on soil were the primary sources of As. carbonarius and As. niger (Leong et al., 2006a, 2006b). Varga et al. (2006) studied the mycobiota of raisins purchased in Hungary. Their results showed that most raisins were heavily contaminated with black aspergilli.
The mycotoxin analysis showed that OTA was found in 70% of the raisin samples used in this study (Table 1). From black and white raisins samples, OTA was detected in 42% and 28% of the total samples, respectively. In comparison, previous UK surveys have reported incidences of OTA in raisins of 85% (MacDonald et al., 1999) and 97% (MAFF, 1999); whereas a German survey (Engel, 2000) reported a 95% overall incidence of OTA in raisins and currants. The OTA was detected in 74% of dried vine fruit samples from Argentinea, and it was detected in 68% and 88% of the black and white fruit (Magnoli et al., 2004). The OTA was present in 67 (79%) of 85 samples of raisins from the Canadian retail market (Lombaert et al., 2004). The percentage of contaminated samples with OTA in our study is similar to the percentages observed in other European countries including Greece, France, Finland, Germany, and the Czech Republic (Miraglia and Brera, 2002; Ostry et al., 2002; Stefanaki et al., 2003). Also, Palumbo et al. (2011) reported that OTA was detected in 93% of the raisin samples collected from California.
As. carbonarius (31.40%), As. niger (11.59%), and As. ochraceus (1.83%) were potential producers for OTA among the mycobiota isolated in this study. Fourteen isolates of As. carbonarius (73.7% of total As. carbonarius isolates) and 2 isolates of As. niger (22.2% of As. niger isolates) were ochratoxingenic isolates (Table 3). Eighty-eight (96.7%) As. carbonarius isolates and one (0.6%) As. niger var. niger isolate from dried vine fruits from the Spanish market were found to be OTA producers (Abarca et al., 2003). From the As. carbonarius strains isolated from grapes in Europe and Israel, 96.7% were OTA producers (Bau et al., 2005). A very small percentage of As. niger aggregate isolated from grapes and sun-dried grapes samples in Spain showed being OTA producers (1.4%), whereas OTA production was detected in 72.6% of As. carbonarius isolates (Valero et al., 2005).
Detection limit was 0.5 μg/kg.
OTA, Ochratoxin A.
As. carbonarius molecular characterization and OTA production
Nineteen strains of As. carbonarius were used to study the genetic diversity among As. carbonarius population isolated from raisin samples. The RAPD-PCR technique was employed using two different primers OPX7 and V6 (Figs. 1 and 2). All carbonarius isolates were repeatedly tested. The polymorphic fragment patterns were reproducible with slight variation in intensity and occasionally in the banding pattern. With the used primers, each isolate yielded 8 to 16 bands. Therefore, under each primer, all amplification products were scored for each isolate according to molecular weight. The band pattern obtained from agarose gel electrophoresis from each primer was digitalized by hand to a two-discretecharacter- matrix (0 and 1 for absence and presence of RAPD bands, respectively), and the data of two primers were combined. The analysis data was based on the Nei and Li coefficient (Nei and Li, 1979). A dendrogram was constructed by the Unweighted paired group method of arithmetic average based on Jaccard's similarity coefficient. The dengrogram constructed from the combined results of two primers (Fig. 3) showed that the percentage of similarity between tested isolates fluctuated from 45% to 87%. This results support that this population has high genetic diversity. Ochratoxigenic isolates (KAUAcar 2, KAUAcar 4, KAUAcar 10, KAUAcar 16, and KAUAcar 19) were scattered in all clusters of the dendrogram. So, the constructed dendrogram showed no correlation between DNA banding patterns and the ability of toxin production. Similar results have been described in other studies with Aspergillus spp., showing no correlation between DNA band profiles and mycotoxins production (Lourenço et al., 2007; Gashgari et al., 2010). Fungaro et al. (2004) compared RAPD patterns of toxigenic and non-toxigenic strains of As. carbonarius isolated from coffee beans. Although the PCR-based assay described by the authors was successfully employed to detect As. carbonarius in coffee samples, no association was found between the RAPD genotype and the ability to produce OTA of the strains. Since the RAPD-PCR technique amplified random fragments of the fungal genome, the fragment that containd gene regulating toxin production may not be amplified using this technique with the used primers as recorded by Gashgari et al. (2010).
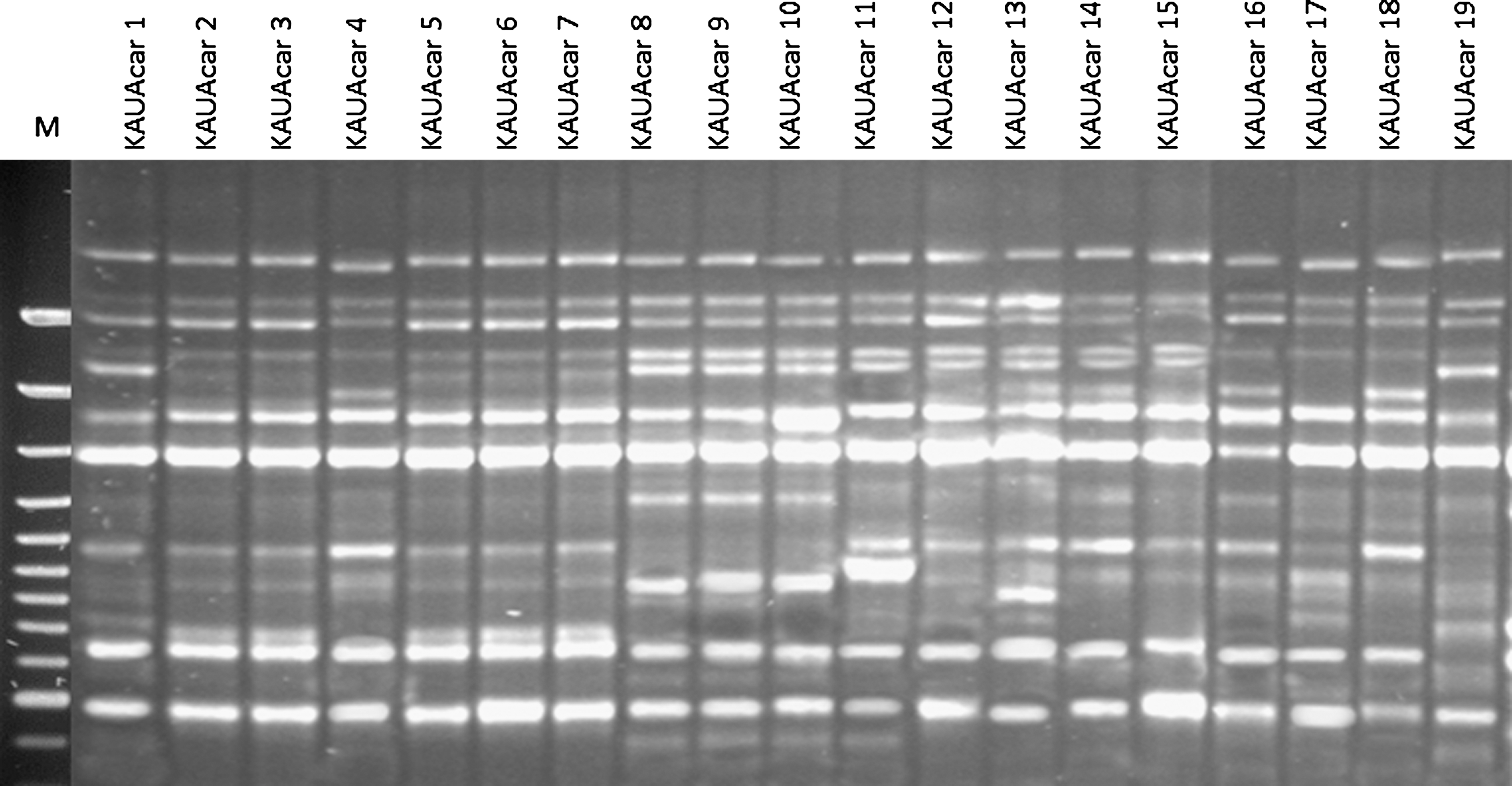
DNA banding patterns from random amplified polymorphic DNA analysis of Aspergillus carbonarius isolates primed by OPX7. Lane M is a 100 bp DNA ladder.
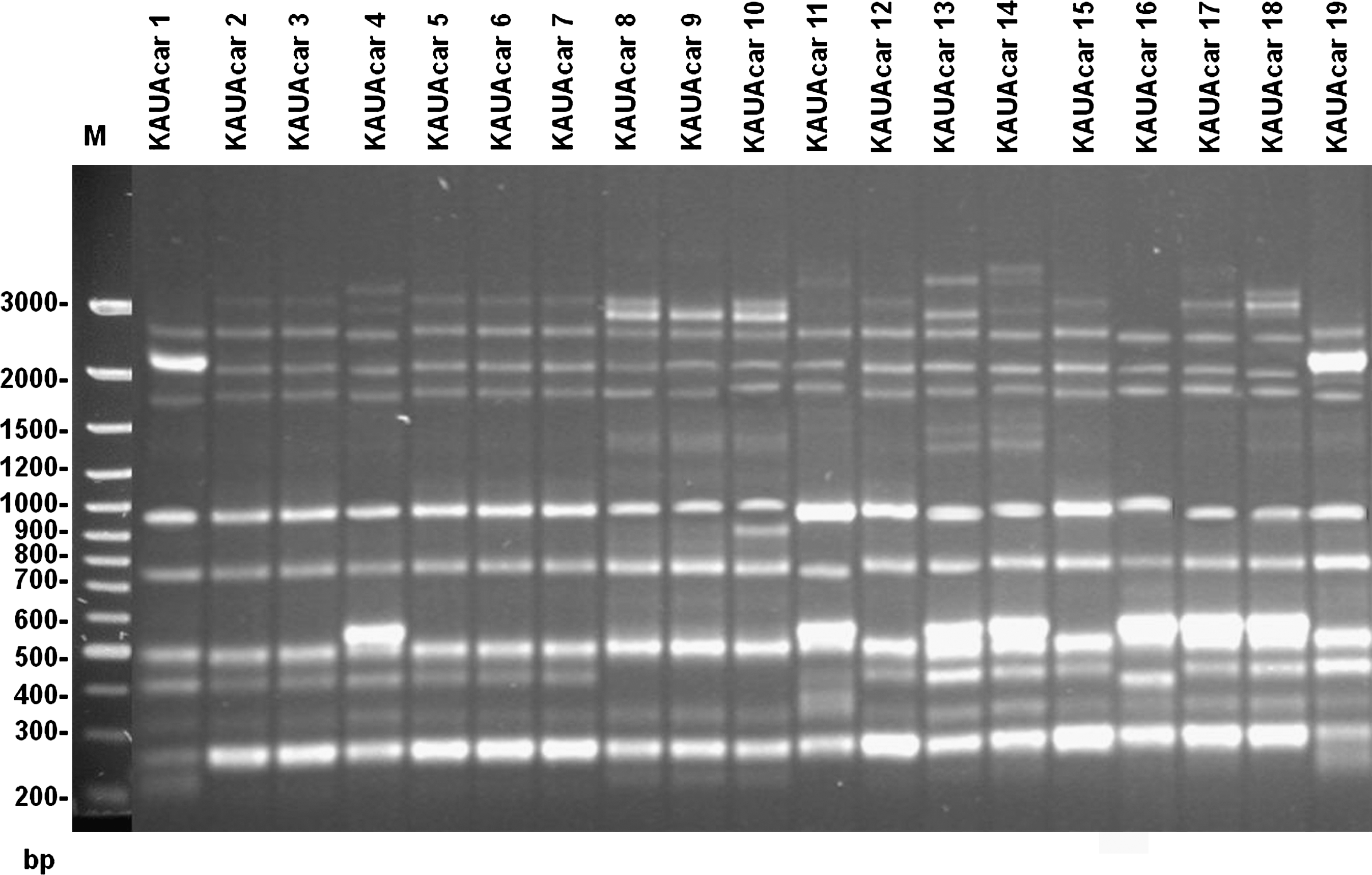
DNA banding patterns from random amplified polymorphic DNA analysis of As. carbonarius isolates primed by V6. Lane M is a 100 bp DNA ladder.
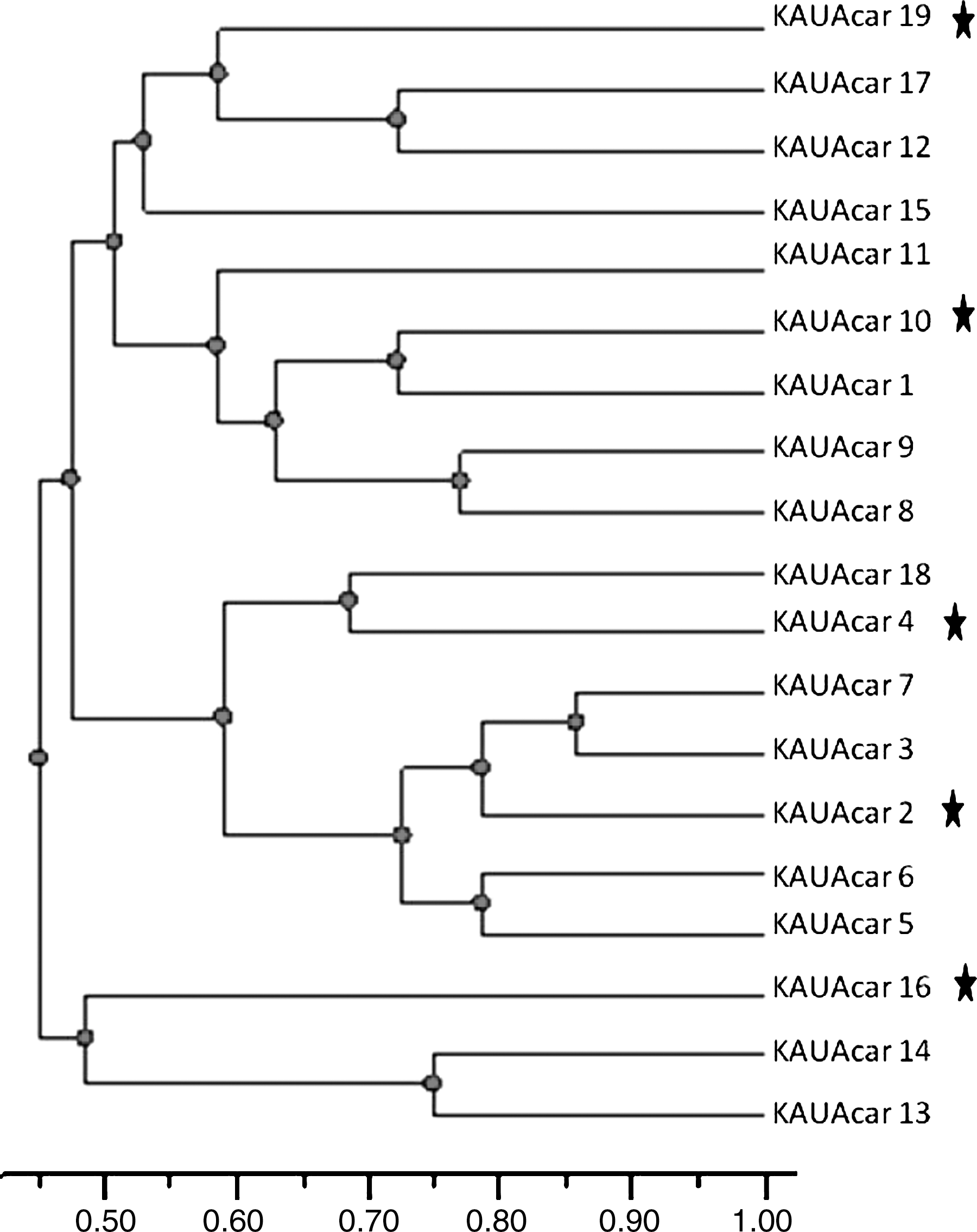
Dendrogram showing relationships among 19 isolates of As. carbonarius isolated from different raisin samples. Genetic distances were obtained by random amplified polymorphic DNA analysis using combining results of OPX7 and V6 primers. Nonochratoxigenic isolates were marked with stars.
Under PCR conditions adopted in this study, the primer pair designed by Sartori et al. (2006) yielded a product of 372 bp from all As. niger strains tested. No amplified product was observed in a reaction using DNA from As. carbonarius, As. Ochraceus, or A. flavus (Fig. 4A). Using the primer pair OPX7F372 and OPX7R372 (Fungaro et al., 2004), all As. carbonarius strains tested yielded a product of 809 bp. Also, no amplified product was observed in a reaction using DNA from As. niger, As. Ochraceus, or A. flavus (Fig. 4B). The PCR assays for As. carbonarius and As. niger identification in pure culture were also successfully applied for detecting an amplicon of 809 and 372 pb, respectively, when using DNA from inoculated raisins (Fig. 5).
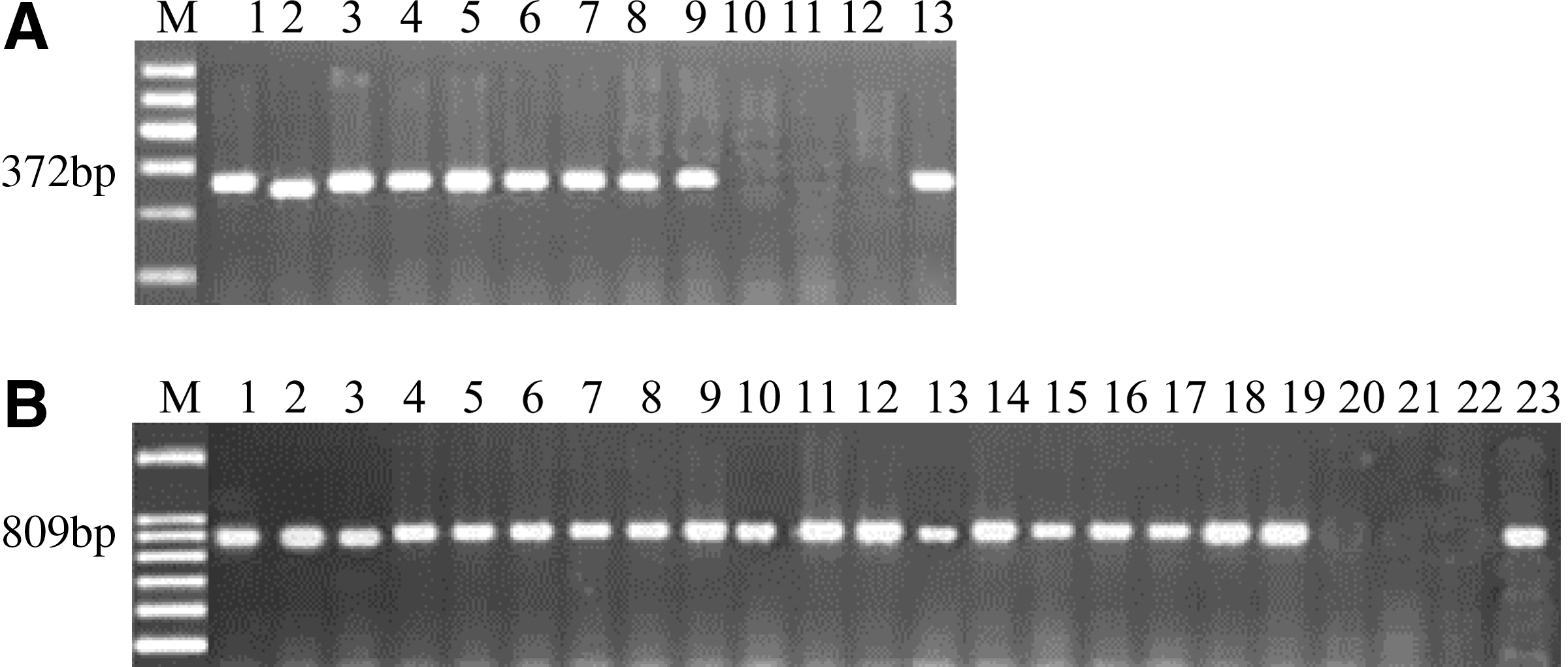
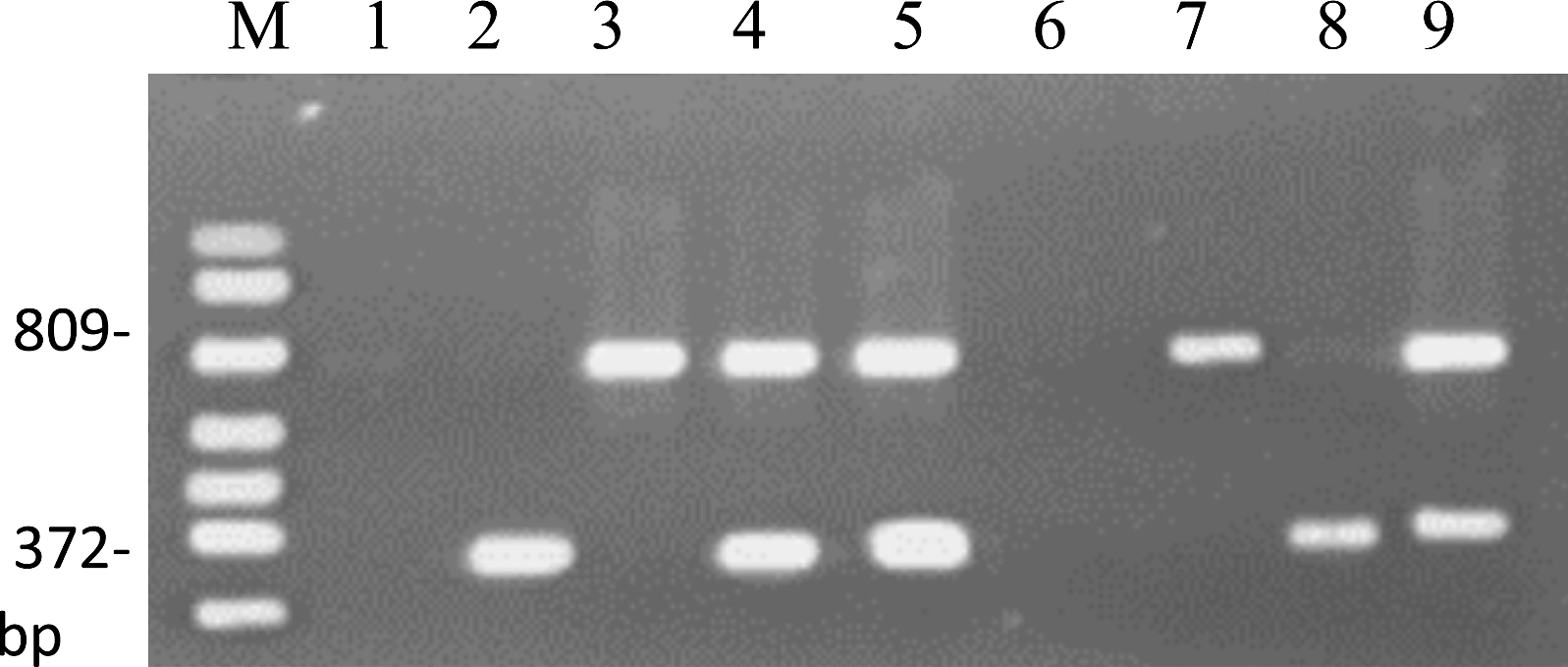
Amplification products obtained by using DNA from inoculated raisins. Lane 1: DNA marker; lane 2: negative control; lane 3: DNA from raisins inoculated with As. niger by using OPX7F372 and OPX7R372 primers; lane 4: DNA from raisins inoculated with As. carbonarius by using OPX7F809 and OPX7R809 primers; lane 5: multiplex PCR using DNA from raisins inoculated with As. carbonarius and As. niger and the 2 sets of primer pairs; lane 6: multiplex PCR using DNA from type strains (TUAniger and TUAcar). Multiplex PCR obtained from raisin naturally contaminated: lane 7: no fungi detection; lane 8: detection of As. carbonarius; lane 9: detection of As. niger; lane 10: detection of As. carbonarius and As. niger.
The multiplex PCR assay for detecting As. carbonarius and As. niger simultaneously was first analyzed using DNA from raisins inoculated with these fungal species. Figure 5 shows the amplification profiles using, simultaneously, the primer pairs designed for As. carbonarius (Fungaro et al., 2004) and As. niger (Sartori et al., 2006). Amplification products of 372 and 809 bp in a single PCR reaction confirmed the presence of As. niger and As. carbonarius, respectively. The applicability of the multiplex PCR assay in quality control of raisins was also analyzed using naturally contaminated samples. This method was previously used by Sartori et al. (2006) for detection of potential ochratoxin-producing Aspergillus species in coffee beans. As shown in Figure 5, this methodology successfully allowed the detection of amplification products from naturally occurring fungi in raisins. The method described in this study represents a much quicker and more reliable detection procedure for the presence of ochratoxigenic fungi found in raisins.
Footnotes
Acknowledgment
This work was supported by a grant (Contract No. 18-011-430) from King Abdulaziz University, Kingdom of Saudi Arabia. The authors are very grateful for the deanship of scientific research at King Abdulaziz and for the financial support they received.
Disclosure Statement
No competing financial interests exist.