
Abstract
Autoinducer-2 (AI-2) is a small molecule that is involved in bacterial cell-to-cell signaling whose precursor formation is mediated by luxS. A luxS mutant of Salmonella Typhimurium PJ002 (ΔluxS) was grown in glucose-containing M-9 minimal medium supplemented with varying concentrations (1×, 10×, and 100×) of long-chain fatty acids (linoleic acid, oleic acid, palmitic acid, and stearic acid) to study the influence of fatty acids on growth rate and macrophage invasion. Additionally, in vitro synthesized AI-2 was added to this medium to identify the influence of AI-2 on S. Typhimurium PJ002 (ΔluxS) growth rate and macrophage invasion. The growth rate constant (k) for each experimental treatment was determined based on OD600 values recorded during 12 h of incubation. There was a significant (p=0.01) increase in the growth rate of S. Typhimurium PJ002 (ΔluxS) in the presence of AI-2 when compared to the phosphate-buffered saline (PBS) control. However, fatty acids either singly or in a mixture were unable to influence AI-2's effect on growth rate. The presence of AI-2 significantly (p=0.02) decreased the invasiveness of S. Typhimurium PJ002 (ΔluxS) towards the murine macrophage cell line, RAW 264.7. However, the fatty acid mixture was able to reverse this reduction and restore invasiveness to background levels. These results suggest that, while AI-2 may enhance the growth rate and reduce macrophage invasion by the luxS mutant S. Typhimurium PJ002 (ΔluxS), fatty acids may influence the virulence in S. Typhimurium (PJ002) by modulating AI-2 activity.
Introduction
Bacterial cells sense signaling molecules produced by other bacterial cells. A common signaling molecule is autoinducer-2 (AI-2), which is produced by both Gram-positive and Gram-negative bacteria (Surette and Bassler, 1998; Surette et al., 1999; Schauder et al., 2001; Winzer et al., 2003; Ahmer, 2004). Autoinducer-2 is produced as a byproduct of the activated Methyl Cycle, in which S-adenosyl-methionine is formed to serve as a methyl donor in cellular metabolism (Chen et al., 2002; Winzer et al., 2003; Miller et al., 2004; Sun et al., 2004; Vendeville et al., 2005).
Studies have shown that AI-2 is a signaling molecule that plays a major role regulating cellular processes in Salmonella, including metabolism, biofilm formation, transport mechanisms, and signal transduction (Sperandio et al., 2001; Ahmer, 2004; Soni et al., 2008; Karavolos et al., 2008; Jesudhasan et al., 2010). In addition, the luxS gene in S. Typhimurium (SL1344) is reported to be essential for the expression of the Salmonella Pathogenicity Island-1 (SPI-1) genes (Choi et al., 2007). However, Perrett et al (2009) who used the same strain reported that the luxS gene did not play a significant role in the expression of SPI-1 genes in S. Typhimurium.
Brominated furanones are known to interfere with AI-2 signaling (Ren et al., 2001, 2004; Widmer et al., 2007a; Girennavar et al., 2008; Vikram et al., 2011). We have shown that fatty acids found in poultry meat and ground beef, such as linoleic acid, palmitic acid, and stearic acid, are also able to inhibit AI-2 activity (Widmer et al., 2007b; Soni et al., 2008). Fatty acids have been previously reported to influence virulence in Salmonella (El-Gedaily et al., 1997; Lucas et al., 2000; Lawhon et al., 2002; Van Immerseel et al., 2003). The hypothesis of this study was that fatty acids will affect the growth rate of S. Typhimurium and influence macrophage invasion. The objective of this study was to understand the influence that fatty acids such as palmitic acid, stearic acid, oleic acid, and linoleic acids have on the growth rate and macrophage invasion abilities of S. Typhimurium in the presence and absence of AI-2 molecules.
Methods
Bacterial strains and cell culture lines
Vibrio harveyi BB170 (a gift from Dr. B. Bassler, Princeton University) was used as the biosensor strain to detect AI-2 activity. Salmonella Typhimurium (isolate no. 87-26254) was obtained from the National Veterinary Service Laboratory (Ames, IA). An isogenic luxS mutant of this strain, designated PJ002 (ΔluxS), was developed in our laboratory as detailed previously (Widmer et al., 2007b; Jesudhasan et al., 2010). The commercially available murine macrophage cell line, RAW264.7 (ATCC no. TIB-71; ATCC, Manassas, VA) was used in the infection assays.
In vitro synthesis of autoinducer-2 (AI-2)
AI-2 was synthesized in vitro according to the published protocol (Sperandio et al,. 2003). The AI-2 activity was confirmed using V. harveyi BB170 as described previously (Lu et al., 2004). For this study, a final concentration of 25 μM of in vitro synthesized AI-2 was used based on a previous study (Soni et al., 2008).
Influence of fatty acid on S. Typhimurium growth rate
Linoleic acid, oleic acid, palmitic acid, and stearic acid were obtained commercially (Sigma-Aldrich, St. Louis, MO). Solutions of linoleic acid, oleic acid, palmitic acid, and stearic acid were prepared in water at concentrations of 0.014, 0.033, 0.02, and 0.0046 mM, respectively. Poultry wash contains these fatty acids at these concentrations (Widmer et al., 2007a). For this study, we used these fatty acids (combined and singly) at 1×, 10×, and 100× of the above concentrations.
The cell pellet from an overnight culture of Salmonella PJ002 (ΔluxS) grown in Luria broth (LB) was washed three times in phosphate-buffered saline (PBS). The washed cells (∼106 CFU) were added to 80 μL of M9 medium (supplemented with 0.2% glucose as the carbon source) in a 96-well clear flat bottom plate (Corning Inc., Corning, NY). Then, either (i) 20 μL PBS (PBS control), (ii) 10 μL AI-2+10 μL of a fatty acid (FA) at a defined concentration (either 1×, 10×, or 100×), (iii) 10 μL PBS+10 μL FA (either 1×, 10×, or 100×), or (iv) 10 μL AI-2+10 μL PBS (AI-2 control) was added to bring the final concentration in each well to 100 μL. The 10-μL aliquots of AI-2 contained 25 μM of AI-2 which was sufficient to elicit clear biosensor strain responses. Each sample was replicated eight times on the plate to verify reproducibility of the observations. Additionally, un-inoculated controls were prepared (20 μL 1×PBS+80 μL M9 medium) to provide a baseline for OD600 readings. The plates were incubated at 37°C for 12 h with moderate shaking in a Tecan Spectrafluor Plus plate reader (Tecan Systems Inc., San Jose, CA). The optical density of each well was measured at a wavelength of 600 nm at 15-min intervals, and the values were automatically recorded.
Cell numbers were estimated using OD600 values (Thibessard et al., 2002; Hung and Mekalanos, 2005; Isaac et al., 2005). The OD600 values were corrected against the un-inoculated control, and then log-transformed and graphed. Time points were selected at the extreme endpoints, encompassing the entire linear portion of the graphed data. These points were then used to calculate the growth rate constant defined as: k=(ln(OD2) – ln(OD1))/(T2 – T1), where OD1 and OD2 were the OD600 values at time 1 (T1) and time 2 (T2), respectively (Isaac et al., 2005). The resulting growth rate constant (k) was used as the basis to compare the effects of AI-2 and the fatty acids. Additionally, to determine if there was any difference in the growth rates of the luxS mutant as compared to its isogenic wild-type, an additional experiment was conducted. Overnight, washed cultures were prepared from the luxS mutant and wild-type strain as described above and inoculated in 80 μL M9 medium+20 μL PBS and incubated at 37°C for 12 h.
Macrophage invasion assay
The RAW264.7 cells were maintained on modified Eagle medium with Earle's modified salts (MEME) and supplemented with 2 mM glutamine and 10% fetal bovine serum (FBS). Six-well plates were used to culture RAW cells to confluence at 37°C in 5% CO2. Overnight cultures (∼109 cfu/mL) of S. Typhimurium wild-type and S. Typhimurium PJ002 (ΔluxS) grown in LB broth were centrifuged (3,500×g for 20 min), and the cell pellets were re-suspended in 400 μL MEME medium (without FBS) individually. S. Typhimurium wild-type and PJ002 (ΔluxS) were used to compare the differences in infectivity rate. The PJ002 (ΔluxS) strain alone was used in the following treatments: either (i) 100 μL PBS (PBS treatment), (ii) 50 μL PBS+50 μL AI-2 (AI-2 treatment), (iii) 50 μL AI-2+50 μL 100X of the combined fatty acids (100× FA+AI-2), or (iv) 50 μL PBS+50 μL 100× of the combined fatty acids (fatty acid alone). The final volume was 500 μL.
Macrophage invasion assays were performed based on published protocols (Sha et al., 2004; Chopra et al., 2006). Bacterial cell suspensions (500 μL), containing approximately 107 bacterial cells were added to the wells and allowed to incubate for 1 h at 37°C in 5% CO2. Based on the counts of RAW cells and bacterial cells, the ratio of macrophage cells to bacteria (MOI) was approximately 1:100. The wells were washed twice with PBS, and 300 μL MEM supplemented with ceftriaxone (100 μg/mL) and 10% heat-inactivated FBS was added to each well. The plates were incubated for 2 h at 37°C in 5% CO2. After incubation, the wells were washed three times with PBS and 400 μL 1% Triton X-100 was added. The plates were incubated at room temperature for 10 min, followed by the addition of 600 μL LB broth to each well. Aliquots were removed, serially diluted in PBS, plated onto LB agar plates, and colonies enumerated after 37°C overnight incubation. The percent infection was calculated as the number of recovered cells (CFU/mL) post-infection divided by the initial inoculum multiplied by 100 (Fahlen et al., 2000; Raffatellu et al., 2005). These assays were repeated twice, with three replications of each treatment. Negative controls (un-inoculated wells seeded with RAW cells) and controls to demonstrate the bactericidal effectiveness of ceftriaxone (Salmonella cell suspension in MEME) were also included. Both the mutant strain and the wild-type strain were used in infection assays where the cell suspensions were prepared in 400 μL MEM medium with 100 μL PBS, as described previously.
Statistical analysis
Statistical analysis was performed using SPSS 11.0 (SPSS, Chicago, IL) by employing either an independent t-test, or when applicable, an analysis of variance (ANOVA) with Dunnitt's test used for post-hoc analysis. Results were considered statistically significant at a calculated p value of ≤0.05.
Results
Influence of AI-2 and fatty acids on S. Typhimurium growth rate
In vitro synthesized AI-2 used in this study showed relative activity of ≈200-fold relative light units (RLU) as compared to a control using the V. harveyi reporter strain (data not included). The mean growth rate (k) of the wild-type and mutant cells was 0.0047 (± 0.0005) and 0.0051 (± 0.0002), respectively. When the mean cell densities (OD600) were compared, treatments with AI-2 had a pronounced increase in cell density compared to treatments that did not contain AI-2 (Table 1). There was a statistically significant (p=0.01) difference in the growth rate of S. Typhimurium in the presence and absence of AI-2 (Table 1).
Significant difference in the mean values of k compared to the phosphate-buffered saline (PBS) control (p=0.01).
The data is based on n=39 replicate samples.
The fatty acid treatments were compared to the PBS control to determine if the fatty acids influenced S. Typhimurium PJ002 (ΔluxS) growth rate (Table 2). Statistical analyses showed that with the exception of linoleic acid at 100×, none of the other fatty acids had any effect on the growth rate of S. Typhimurium PJ002 (ΔluxS) (Table 2). The combined fatty acid mixture also did not have any significant effect on S. Typhimurium PJ002 (ΔluxS) growth rate (Table 2). The influence of fatty acids on S. Typhimurium PJ002 (ΔluxS) growth rate in the presence of AI-2 was investigated (Table 3). A one-sided Dunnett's test was used for statistical analysis. None of the k values showed any statistically significant difference (p>0.05) for the combined FA+AI-2 treatments compared to the AI-2 control, suggesting that the fatty acids did not influence the AI-2's effect on PJ002 (ΔluxS) growth rate (Table 3).
Significant difference in the mean values of k compared to the phosphate-buffered saline (PBS) control (p=0.05).
Treatment: fatty acids singly, or combined, at a nominal (1×), 10-fold (10×), or 100-fold (100×) concentration. PBS indicates M9 medium supplemented with PBS.
Data based on n=7 replicate samples.
Treatment: fatty acids singly, or combined, at a nominal (1×+AI-2), 10-fold (10×+AI-2), or 100-fold (100×+AI-2) concentration. AI-2 indicates M9 medium supplemented with AI-2.
Data based on n=7 replicate samples.
Influence of AI-2 and fatty acids on macrophage invasion by S. Typhimurium
Though the luxS mutant appeared to exhibit a slight increase in its macrophage infection efficiency as compared to the wild-type (1.4% and 0.19%, respectively) (Fig. 1), this was not statistically significant (p=0.2). In the presence of AI-2, however, the infection efficiency of Salmonella Typhimurium luxS mutant decreased as compared to the PBS control (Fig. 2). The presence of AI-2 in the sample resulted in a statistically significant (p=0.02) reduction in infection efficiency (0.3%) compared to the PBS control (0.76%). However, when the combined FA mixture was added to the AI-2–containing sample, S. Typhimurium's infection efficiency increased to 0.49% (Fig. 2). There was no statistical difference (p=0.13) in the infection efficiency between the FA+AI-2 treatment and the PBS control treatment. To further investigate this influence of FA, studies were performed to determine if the combined fatty acid treatment could improve infection efficiency. The 100× fatty acid (FA) treatment did appear to have an improved infection (1.54%) compared to the AI-2 treatment (p=0.01). However, there was no statistically significant difference (p>0.05) in the percentage infection between the FA mixture and PBS, and the FA+AI-2 treatments (Fig. 2).
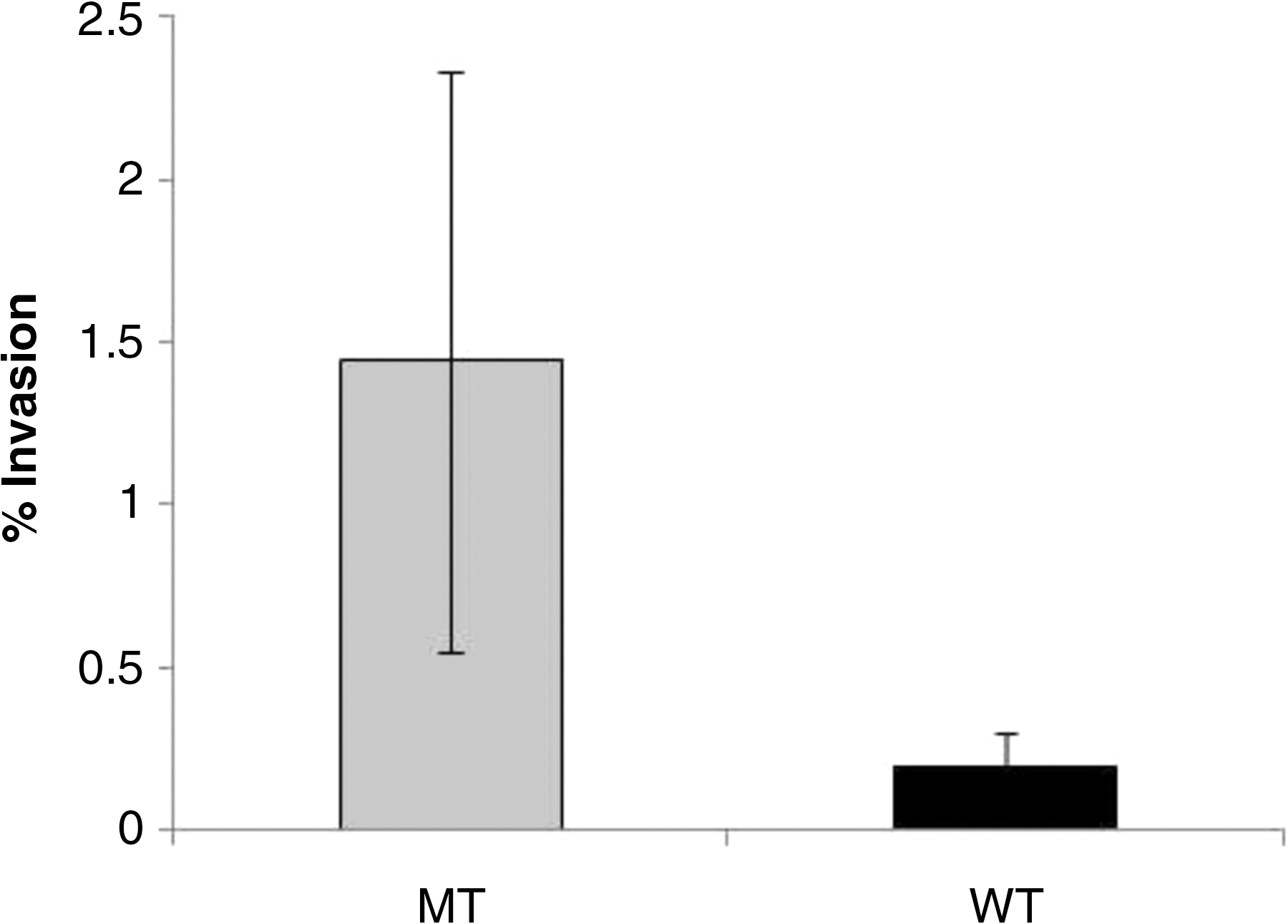
Infection efficiency (% invasion) of Salmonella Typhimurium wild-type (WT) and luxS mutant (PJ002 ΔluxS) (MT) in macrophage cells. The error bars represent the means for each treatment (n=6)±standard error.

Infection efficiency (% invasion) of Salmonella Typhimurium luxS mutant (PJ002) in the presence of autoinducer AI-2, 100× fatty acid mixture (FA), 100× fatty acid mixture combined with AI-2 (FA+AI-2), or phosphate-buffered saline (PBS; control) in macrophage cells. The error bars represent the means (± SE) for each treatment based on six replicate samples. Experimental treatments with different superscripts denote statistically significant differences in percentage invasion as compared to the PBS control. The asterisk (*) represents statistically significant differences in the percentage invasion between the FA and AI-2 experimental treatments.
Discussion
Calculating the growth rate constant (k) based on the linear portion of the exponential growth phase is a useful tool for understanding the growth kinetics of bacteria (Zwietering et al., 1990; Chrzanowski et al., 1993). This approach has been used previously to compare the growth and survival of bacteria in foods under varying conditions (Bachrouri et al., 2002; Mamani et al., 2003). There was no significant difference in growth rate of AI-2–deficient PJ002 (ΔluxS) and the wild-type AI-2–producing cells in M9 medium. The growth rate (k) of the wild type and mutant cells were 0.0047 (± 0.0005) and 0.0051 (± 0.0002), respectively, suggesting no significant differences in their growth rate in M9 minimal medium. However, S. Typhimurium PJ002 (ΔluxS) does respond to the addition of AI-2 to the M9 minimal medium with an increased growth rate (Table 1). The luxS mutant strain PJ002 (ΔluxS) responded with an increased growth rate because of the supplemented (25 μM) in vitro synthesized AI-2, which was not present in the PBS buffer control (Table 1). The lack of detectable difference in the growth rate of the wild-type and luxS mutant in M9 medium could be due to the low levels of AI-2 produced by the wild-type cells in M9 minimal medium (which contained only 0.2% glucose). These concentrations were possibly below a threshold to elicit an enhanced growth response. The AI-2 activity produced by the wild-type S. Typhimurium in M9 medium was only one fourth of the activity produced by in vitro synthesized AI-2 (data not included). Other investigators often employed rich media such as LB or LB supplemented with 0.5% glucose to demonstrate that AI-2 influences bacterial growth (Surette and Bassler, 1998; Surette et al., 1999; Taga et al., 2003). The production of AI-2 is promoted in media characterized by high nutrient content, low pH, and high osmolarity (De Lisa et al., 2001; Surette et al., 1999).
We had previously reported that fatty acids such as palmitic acid, stearic acid, oleic acid, and linoleic acid inhibited AI-2 activity 25–99% (Widmer et al., 2007a; Soni et al., 2008). Since we had observed that AI-2 enhanced the growth rate of PJ002 (ΔluxS) in M9 medium, we hypothesized that the addition of fatty acids would inhibit the growth rate stimulation by AI-2. However, the addition of various fatty acids to M9 medium in varying concentrations (singly or in combination) did not influence growth rate (as compared to the PBS control) (Table 2). Only one fatty acid (linoleic acid) at 100× concentration had a statistically significant effect (p=0.05) on growth rate compared to the PBS control. The fatty acids either singly or in combination did not appear to influence the growth rate of PJ002 (ΔluxS) in M9 medium in the presence of AI-2 (Table 3). These results imply that in M9 minimal medium the inhibitory effects of fatty acids on AI-2 are not pronounced.
The ability of Salmonella to evade host defenses by surviving in macrophages is a key aspect of determining the efficacy and progression of Salmonella infection (Rosenberger et al., 2000). The ability to not only survive phagocytosis, but also replicate within macrophage vacuoles is a key aspect for the pathogenicity of Salmonella (Richter-Dahlfors et al., 1997; Garvis et al., 2001). This study demonstrates that AI-2 impedes invasion of Salmonella in murine macrophages, and such reduction in infectivity can be reversed with the addition of fatty acids (Fig. 2). This result agrees with our recent report showing that AI-2 actually down-regulates the expression of the Salmonella Pathogenicity Island-1 (SPI-1) and thereby down-regulates S. Typhimurium virulence and invasion genes (Jesudhasan et al., 2010). It is possible that the enhanced invasion that was observed in the presence of the fatty acids is due to fatty acids modulating or limiting the influence of AI-2 (which would normally down-regulate virulence and invasion).
There are several reports of fatty acids influencing the virulence of Salmonella (El-Gedaily et al., 1997; Lucas et al., 2000; Lawhon et al., 2002; Van Immerseel et al., 2003). Lucas et al. (2000) found that genes responsible for the metabolism and uptake of fatty acids affected the expression of hilA, a key gene involved in S. Typhimurium virulence. El-Gedaily et al. (1997) reported that short-chained fatty acids enhanced virulence, while long-chain fatty acids reduced virulence. Studies by Lawhon et al. (2002) demonstrated that proprionate and butyrate reduced in vitro Salmonella Enteritidis infection. However, when combined with other fatty acids (that mimicked the contents and concentrations of an adult chicken caecum), infection was normalized.
Conclusion
This study demonstrates that, in the low nutrient M9 medium, the addition of AI-2 enhances the growth rate in AI-2–deficient, S. Typhimurium PJ002 (ΔluxS). Long-chain fatty acids were unable to reverse this increased growth rate due to AI-2. However, these fatty acids were able to reverse the reduction in macrophage invasion that AI-2 causes in S. Typhimurium PJ002 (ΔluxS). These studies provide evidence that fatty acids and AI-2 may play a role in the virulence of S. Typhimurium.
Footnotes
Acknowledgments
The technical assistance provided by Tara Hans and Martha Cepeda is gratefully acknowledged. This study was supported by Texas AgriLife Research of the Texas A&M University System (Hatch grant H-8708).
Disclosure Statement
No competing financial interests exist.