
Abstract
In this study, we demonstrate that purA and purB transposon mutants of serotype 4b Listeria monocytogenes were severely impaired in their ability to colonize the gastrointestinal tract and cause systemic infection of the spleen, liver, and gallbladder following intragastric inoculation of A/J mice. The mutant strains were also impaired in their ability to multiply within Caco-2 human intestinal epithelial cells. Neither mutant was affected in resistance to synthetic gastric fluid (pH 4.5). These findings indicate that purine biosynthesis is critical for gastrointestinal virulence of L. monocytogenes serotype 4b in mice.
Introduction
L
Purine biosynthesis has been implicated in virulence in several Gram-positive and Gram-negative bacterial pathogens, including Bacillus anthracis (Ivanovics et al., 1968), Yersinia pestis (Brubaker, 1970), Salmonella enterica (McFarland and Stocker, 1987), Streptococcus pneumoniae (Polissi et al., 1998), and Staphylococcus aureus (Lan et al., 2010). In L. monocytogenes, an adenine auxotroph was found to be reduced in virulence (by 1.5 log10) in a murine model utilizing intravenous infection and showed reduced growth rate in macrophage-like J774 cells or the epithelial cell line Henle 407 (Marquis et al., 1993). Mutant screens also showed that mutants with insertions in purine and pyrimidine biosynthesis genes were impaired in their intracellular growth in the intestinal epithelial cell line Caco-2 (Schauer et al., 2010). Adenosine biosynthesis has been shown to be required for growth of L. monocytogenes in porcine bile (Dowd et al., 2011).
Limited information is currently available on the role of purine biosynthesis in listeriosis following transmission of the pathogen via the gastrointestinal route. Furthermore, previous studies have all utilized strains of serotype 1/2a (10403S or EGDe) (Marquis et al., 1993; Schauer et al., 2010; Dowd et al., 2011). However, serotype 4b strains differ both in serotype and in lineage from those of serotype 1/2a, and involvement of virulence determinants may differ between serotype 1/2a and 4b strains.
In this report, we have employed the previously described A/J mouse model of gastrointestinal listeriosis (Czuprynski et al., 2003a,b) to investigate the virulence of purA and purB mutants of serotype 4b L. monocytogenes. In addition, we have assessed the ability of these purine biosynthesis mutants to grow in Caco-2 cells and to survive in synthetic gastric fluid.
Methods
Bacterial strains and culture conditions
The serotype 4b L. monocytogenes strains employed in the study were F2365 (epidemic clone I, 1985 Mexican style cheese outbreak) (Nelson et al., 2004) and H7550-CdS (epidemic clone II, 1998–1999 hot dog outbreak) (Elhanafi et al., 2010). The purA mariner-based mutant of F2365 and the purB mariner-based mutant of H7550- CdS were designated ROA14 and J22F, respectively. Bacteria were grown in tryptic soy broth (TSB) with 0.7% Yeast Extract (Becton, Dickinson and Co., Sparks, MD) or on trypticase soy agar (TSA) plates containing 1.5% agar (Becton, Dickinson and Co.). Cultures were routinely grown for 20 h at 37°C. For long-term preservation of the strains, we used Listeria-coated beads (Copan Diagnostics Inc., Corona, CA) following the instructions of the manufacturer.
To prepare L. monocytogenes cells for inoculation of mice, cell cultures, and synthetic gastric fluid, a single Listeria-coated bead was inoculated into 50 mL of brain heart infusion broth (BHI) (Becton, Dickinson and Co.) in a 250-mL Erlenmeyer flask. The culture was incubated for 20 h with shaking at 37°C until stationary phase growth was reached. The cells were recovered by centrifugation and resuspended in an equal volume of phosphate-buffered saline (PBS). The optical density of the bacterial suspension was determined, and the colony-forming units (CFU) were extrapolated from a standard curve. Appropriate dilutions of the bacterial suspension were made in sterile PBS to achieve the desired bacterial concentration, which was verified in each experiment by plate counts on TSA.
Construction and characterization of purine biosynthesis mutants
Mariner-based transposon mutant libraries of strains F2365 and H7550-CdS were constructed using plasmid pMC39 (kindly provided by H. Marquis, Cornell University) as previously described (Cao et al., 2007; Azizoglu and Kathariou, 2010). Localization of the transposon insertion by arbitrary polymerase chain reaction (PCR) and determination of transposon copy number by Southern blots with the transposon-specific ermC probe were determined as previously described (Cao et al., 2007; Azizoglu and Kathariou, 2010).
Mouse inoculation experiments
We assessed the virulence of mutant and wild-type strains of L. monocytogenes using a previously described model of gastrointestinal listeriosis (Czuprynski et al., 2003a). Briefly, 5–6-week-old female A/J mice (Harlan Sprague-Dawley, Indianapolis, IN) were housed under microisolator caps at the School of Veterinary Medicine animal care facility for at least 1 week before being used in an experiment. Mice were anesthetized by i.p. injection with sodium pentobarbital (0.75 mg per 25 g body weight) (Abbott, Abbott Park, IL) and then inoculated intragastrically with approximately 1.5×106 CFU of L. monocytogenes (in a total volume of 0.2 mL) via a 1.5-inch 24-gauge stainless steel feeding needle attached to a 1-mL syringe as described previously (Czuprynski et al., 2003b). Three days later (at the peak of the bacterial burden in wild-type mice), the mice were euthanized by asphyxiation with CO2 and exsanguinated by cardiac puncture. Portions of the spleen, liver, cecum, and gallbladder were removed, weighed in sterile weigh boats (except for gallbladder), and placed in sterile tissue grinders that contained 1 mL of cold sterile saline (0.85%). The tissues were homogenized, diluted in sterile saline, and plated in duplicate onto TSA plates or modified Oxoid agar plates (cecal samples). Data are expressed as the mean±SEM log10 CFU of L. monocytogenes per g tissue (wet weight), except for the gallbladder data, which are expressed as CFU/organ, and blood, which is expressed as CFU/mL. The limit of detection was 1.0 log10 CFU. Tissues that did not yield any colonies were assigned a value of 0.95 log10 CFU for calculation of the mean±SEM of that treatment group. This protocol was approved by the University of Washington–Madison School of Veterinary Medicine institutional animal care and use committee.
Caco-2 invasion and intracellular growth assay
The ability of L. monocytogenes to invade and multiply within the human colonic adenocarcinoma cell line Caco-2 (American Type Culture Collection [ATCC] HTB37, Rockville, MD) was determined as described previously (Faith et al., 2005). Caco-2 cells were incubated in 24-well tissue culture plates that contained Dulbecco's modified Eagle's medium (DMEM) supplemented with 25 mM glucose, 1% nonessential amino acids, 2 mM
Survival of L. monocytogenes in synthetic gastric fluid
Synthetic gastric fluid (SGF) was prepared as described previously (Cotter et al., 2001). Briefly, 8.3 g of proteose peptone, 3.5 g of
Statistical analysis
Data were analyzed by a one-way analysis of variance using GraphPad Prism version 4.0 (GraphPad Software, Inc., San Diego, CA). If a significant F value was obtained (p<0.05), the Tukey-Kramer or Mann-Whitney test was performed to determine whether the means of treatment groups differed from those of controls. Statistical significance for all comparisons was set at p<0.05.
Results
Mutants in purA and purB are severely impaired in growth on trypticase soy agar with blood but grow normally on blood-free medium
Screening of 1,906 mariner-based transposon mutants of H7550-CdS for hemolytic activity on blood agar plates revealed that one mutant (J22F) barely grew on this medium. However, the mutant grew as well as wild-type bacteria and other mutants on TSA plates without blood and in liquid medium (Suppl. Fig. S1; Supplementary Material is available online at
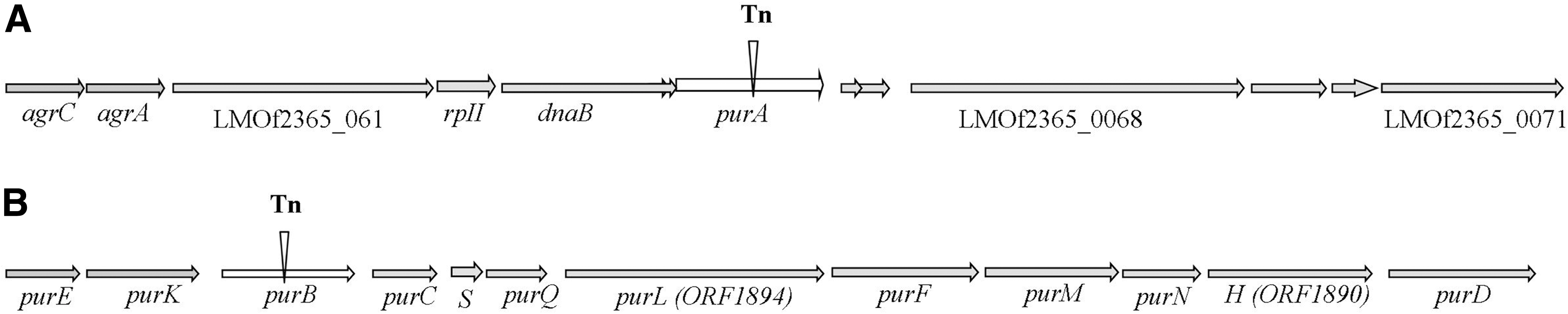
Genomic organization of the regions harboring purA and purB in Listeria monocytogenes. Arrows indicate direction of transcription; vertical black bars indicate transposon insertion.
Mutants in purA and purB are severely attenuated in virulence for intragastrically (i.g.) inoculated mice
The purA mutant ROA14 displayed reduced ability to cause systemic infection in i.g. inoculated mice. Numbers of ROA14 recovered from the liver and spleen were 4–5 log10 CFU less than those of the wild-type parental strain F2365; a significant reduction (2–4 log10) was also noted in CFUs recovered from the cecum, blood, or gallbladder (Fig. 2). The impairment of virulence was even more pronounced with J22F, the mutant in purB. Recovery of this mutant from the liver and spleen following i.g. inoculation was reduced by more than 6 log10 CFU compared with the wild-type parent strain H7550-CdS (Fig. 2). Significant reductions (approximately 3 log10 CFU) were also observed in recovery of the purB mutant from the cecum, blood, and gallbladder of the mice (Fig. 2). Thus, the purA and purB mutants were severely impaired in their ability to cause gastrointestinal or systemic infection following i.g. inoculations. In contrast, the wild-type parental counterparts colonized the gastrointestinal tract and caused a substantial systemic infection that resulted in recovery of large numbers of bacteria from the cecum, spleen, liver, gallbladder, and blood (Fig. 2).
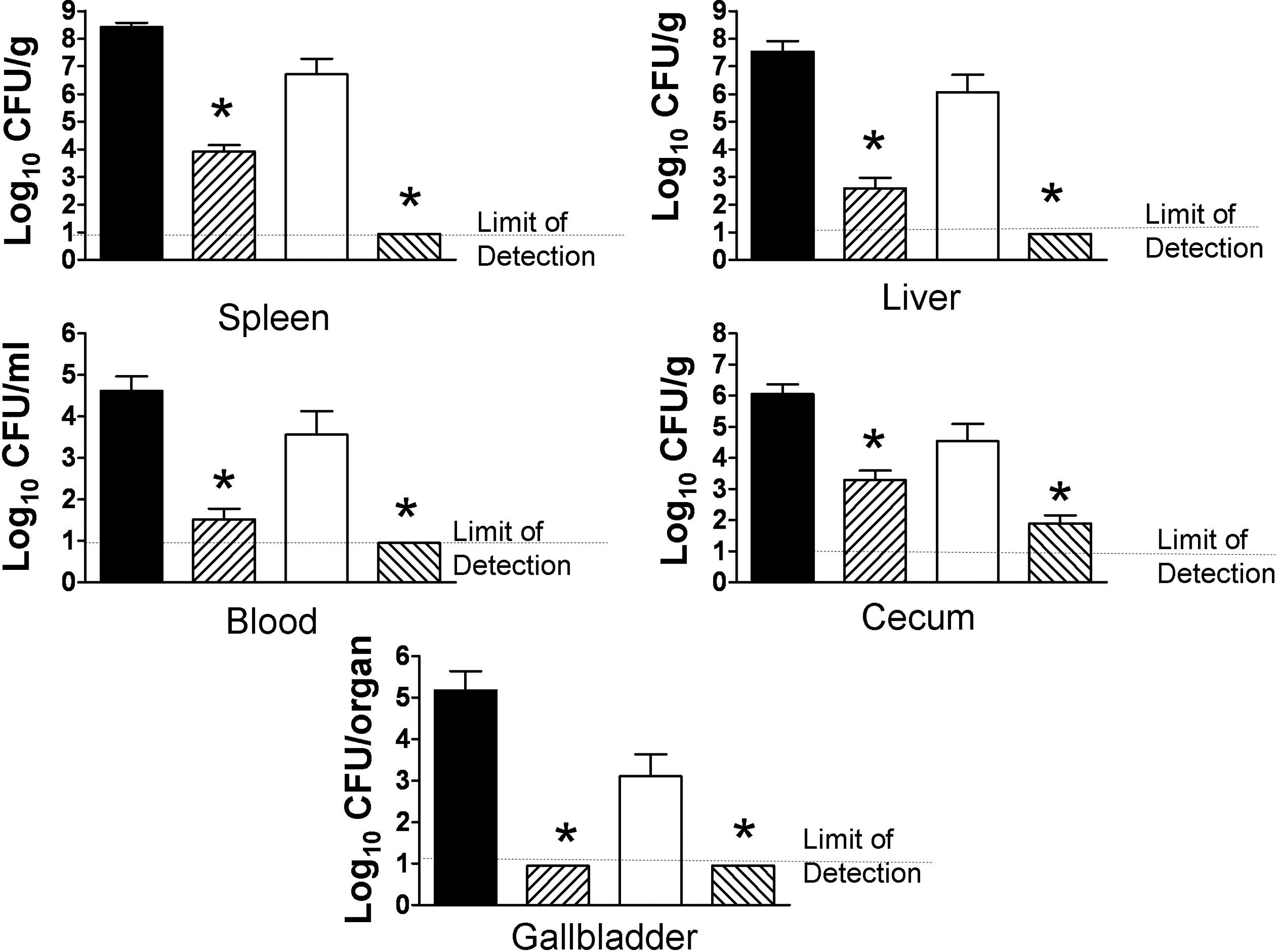
purA and purB mutants of serotype 4b Listeria monocytogenes are severely impaired in ability to cause systemic infection following i.g. inoculation in A/J mice. A/J mice were inoculated i.g. with approximately 1.5×106 CFU of the purA mutant ROA 14 (cross-hatched bars) and its parental wild-type strain F2365 (solid bars; six mice each group) or with the purB mutant J22F (cross-hatched bars) and its parent strain H7550-CdS (open bars; 18 mice each group). Intragastric inoculations and determination of CFU in liver, spleen, cecum, blood, and gallbladder were conducted as described in the Methods. Data are expressed as the mean±SEM log10 CFU/g tissue, mL of blood, or entire organ (gallbladder). Asterisks indicate p<0.05 compared to the wild-type parent.
purA and purB mutants are deficient in intracellular growth in Caco-2 cells
When invasion was assessed, neither the purA nor the purB mutant differed significantly from their respective parental strains. However, invasion by either the purA mutant or its parent strain (F2365) was relatively low compared with strain H7550Cds or its purB mutant (J22F) (Fig. 3). Most importantly, there was a substantial reduction (approximately 3 log10 CFU) in intracellular multiplication of both ROA14 (purA mutant) and J22F (purB mutant) compared to their respective parental strains (Fig. 3).
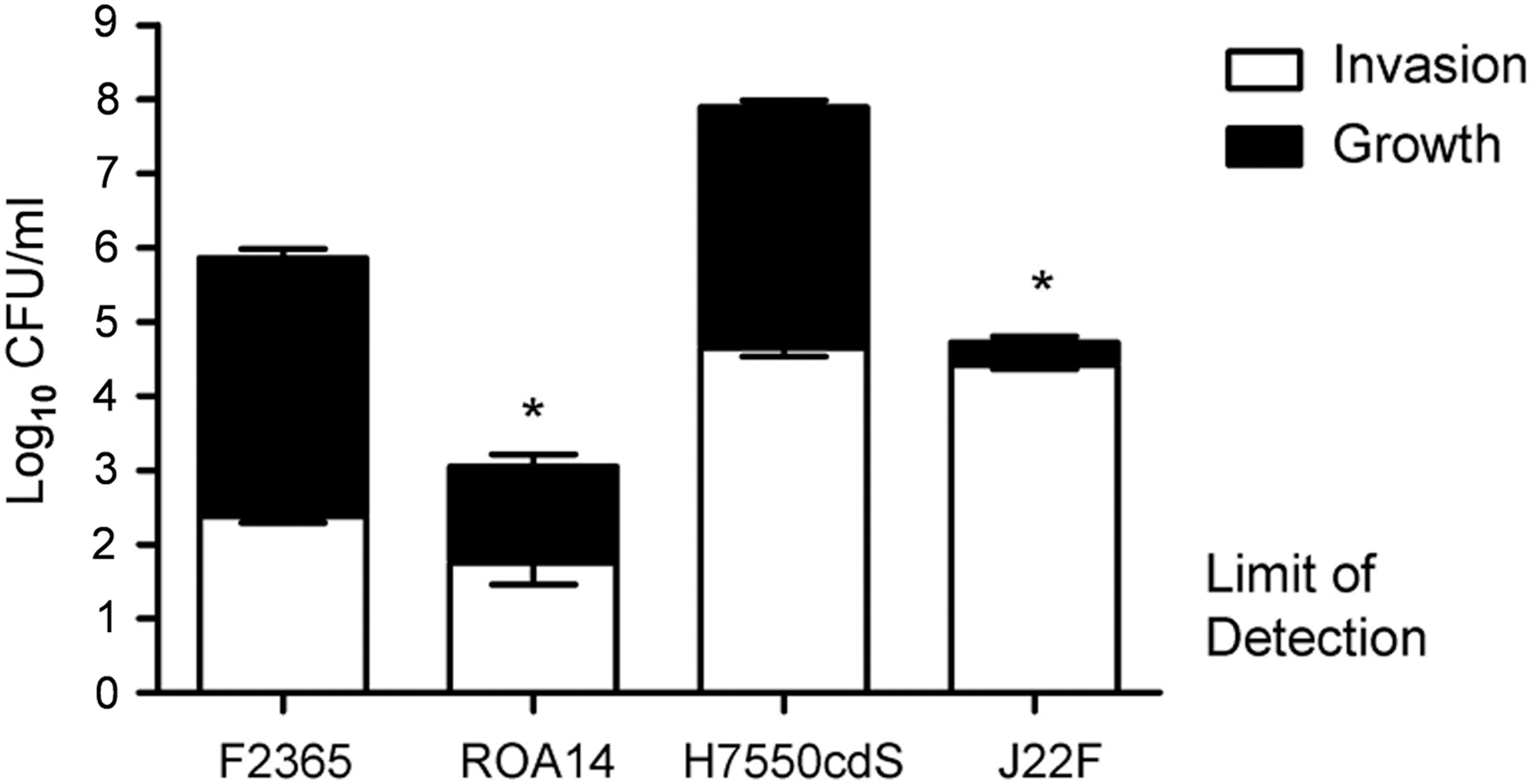
purA (ROA 14) and purB (J22F) mutants of Listeria monocytogenes serotype 4b are severely diminished in their ability to multiply within Caco-2 intestinal epithelial cells. Invasion and intracellular multiplication were assessed as described in the Methods. Briefly, Caco-2 cells were incubated in complete medium for 15–18 days at 37°C. To assess invasion, monolayers were incubated with L. monocytogenes (1×107 CFU/ml, 3 wells per group) for 1 h, washed 5×with Hanks' balanced salts solution (HBSS) and then incubated for an additional 2.5 h in medium with 5 μg/mL gentamicin. Monolayers were washed 5×in phosphate-buffered saline (PBS), lysed in 1% Triton-X-100, and a portion of the lysates (0.25 mL) diluted in PBS and plated in duplicate on trypticase soy agar (TSA) plates. To quantify intracellular multiplication, the infected monolayers were incubated in medium with 5 μg/mL gentamicin (2.5 mL final volume) for an additional 12 h at 37°C before being lysed and the lysates diluted and plated as described above. Data are expressed as the mean±SEM log10 CFU per well of two separate experiments (3 wells per group in each experiment). Invasion data are illustrated by the open portion of the bars; intracellular multiplication by the closed portion of the bars. Asterisks indicate p<0.05 compared to the wild-type parent for intracellular growth; invasion did not differ (p>0.05) between the mutants and their respective parent strains.
purA and purB mutants are not affected in their ability to survive in synthetic gastric fluid
To determine whether purA and purB mutants were impacted in their ability to survive under conditions that mimic those in the upper gastrointestinal tract, we assessed survival of the bacteria in synthetic gastric fluid at pH 4.5. We found no significant impairment in the ability of log-phase cells of either mutant to survive in synthetic gastric fluid as compared with their respective wild-type parent strains (Fig. 4). Survival relative to wild type also was not impacted when stationary phase cells were used (data not shown).
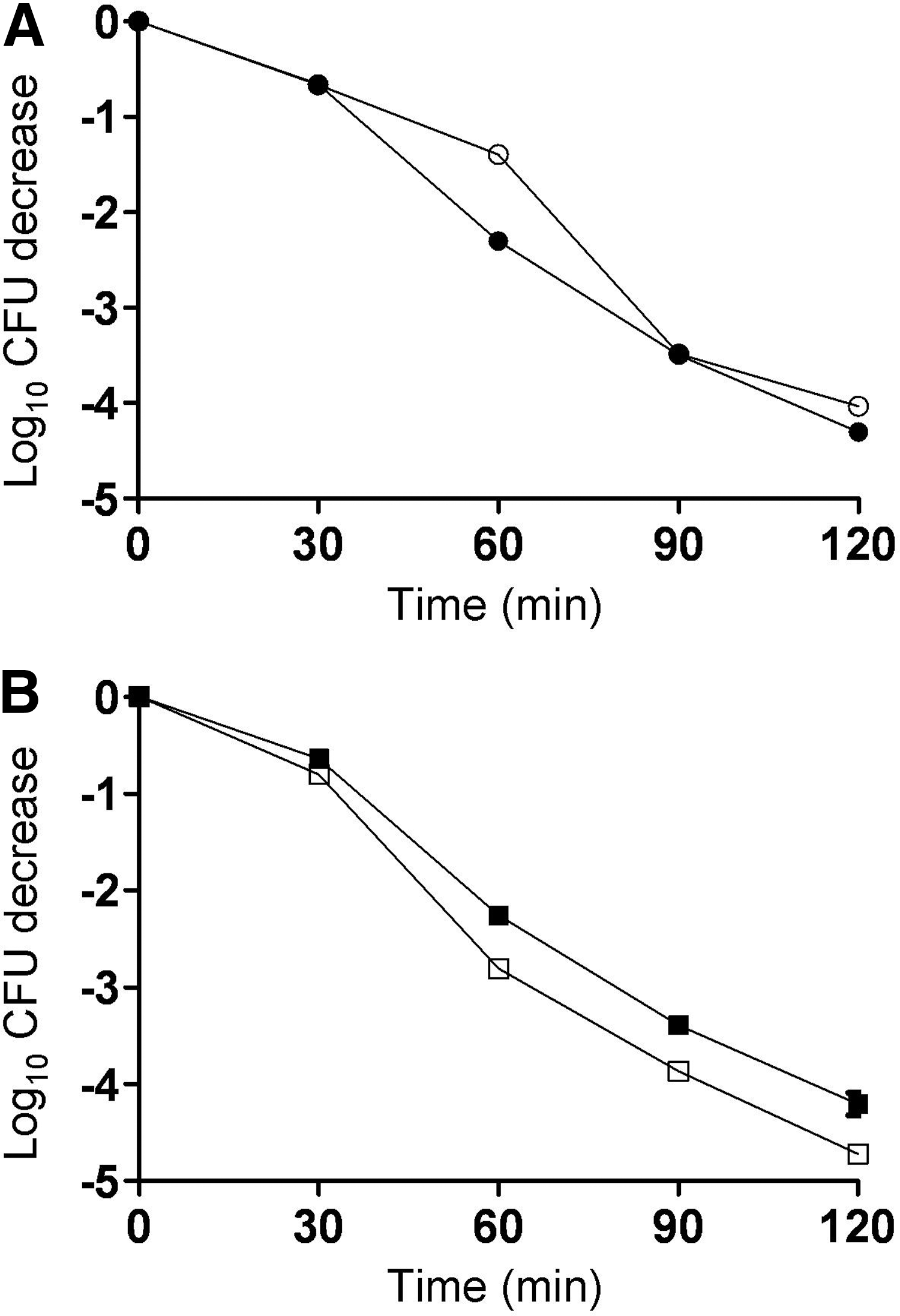
purA and purB mutants of Listeria monocytogenes serotype 4b are not deficient in survival in synthetic gastric fluid (pH 4.5). Synthetic gastric fluid preparation and survival assessments were done as described in the Methods.
Discussion
The results of this study provide evidence that purine biosynthesis is required for virulence of L. monocytogenes inoculated into the gastrointestinal tract of A/J mice. Transposon mutants in purA and purB, constructed in two different serotype 4b parent strains, exhibited severely diminished ability to colonize the gastrointestinal tract and cause systemic infection in i.g. inoculated mice, as compared to mice inoculated with the wild-type parental strains. Furthermore, both mutants were significantly impaired in their ability to grow in Caco-2 cells. Although the purA mutant and its wild-type parent were less invasive for Caco-2 cells than the purB mutant and its wild-type parent, we are reluctant to make inferences regarding the relative invasive abilities of the two pairs of strains. Instead, we focus on the diminished intracellular growth of both the purA and purB mutants. This observation is consistent with findings from a previous report that suggested the importance of purine synthesis for intracellular multiplication of L. monocytogenes in Caco-2 cells (Schauer et al., 2010). It is interesting that the purB mutant is particularly poor at intracellular multiplication in Caco-2 cells. Furthermore, our data revealed a low level of recovery of this mutant from the cecum and nearly complete absence from the liver and spleen.
The identification of similar phenotypes in mutants from two different strains and verification that the insertions were in both cases in purine biosynthesis genes suggest that the decreased virulence phenotypes of the mutants indeed were associated with defects in purine biosynthesis. Polar effects of the transposon insertion are unlikely in the case of purA, which is not part of an operon (Toledo-Arana et al., 2009; Schauer et al., 2010). However, purB is transcribed in a polycistronic message (purD-purE) that includes the other pur genes shown in Fig. 1 (Toledo-Arana et al., 2009). Thus, the purB mutant may have impaired expression of downstream genes that are also involved in purine biosynthesis. In the absence of genetically complemented derivatives of the mutant strains, it is not possible to determine whether the phenotype of this mutant is specific to inactivation of purB, or can be attributed to polar effects on other purine biosynthesis genes downstream of purB.
Virulence attenuation of purine biosynthesis mutants has been demonstrated with several other bacterial pathogens, including Salmonella (McFarland and Stocker, 1987), Yesinia pestis (Brubaker, 1970), Bacillus anthracis (Ivanovics et al., 1968; Jenkins et al., 2011), Streptococcus pneumoniae (Polissi et al., 1998), and Staphylococcus aureus (Lan et al., 2010). These deficits in virulence are not simply the result of a general impairment in bacterial growth, as purine biosynthesis mutants have been shown to grow at a rate similar to wild-type cells in various types of complex media in vitro (Jenkins et al., 2011). Interestingly, the L. monocytogenes purA and purB mutants in this study were identified precisely because of their severely impaired growth on sheep blood agar, while growth on agar media without blood, or in liquid, was not impacted. The findings suggest that in the presence of blood the agar-grown bacteria could not utilize purines exogenously available in the medium, but the underlying mechanisms remain unidentified. The blood-sensitive phenotype may be specific to serotype 4b strains, as it was not reported in previous studies on purine biosynthesis mutants of L. monocytogenes (including mutants in purA and purB), which employed the serotype 1/2a strains 10403S and EGDe (Marquis et al., 1993; Schauer et al., 2010; Dowd et al., 2011).
Another difference between the phenotype of the mutants described here and those in previous studies is the extent of the impairment in virulence in i.g. inoculated mice and in intracellular growth in the Caco-2 intestinal epithelial cell line. The impact of the mutations on virulence was significantly greater in the current study than reported previously for adenine auxotrophs of L. monocytogenes 10403S (Marquis et al., 1993). Possible reasons include the different serotype and lineage of the strains used in the studies, as well as the different routes of infection (intragastric in the current study vs. intravenous in the study by Marquis et al. [1993]). The impact on virulence observed using the i.g. route and serotype 4b strains of L. monocytogenes was clearly greater than that reported for serotype 1/2a L. monocytogenes (Marquis et al., 1993) but approximated that observed with other bacteria, such as Salmonella and Yersinia pestis (Brubaker, 1983; McFarland and Stocker, 1987).
The purine biosynthesis mutants analyzed here also displayed lower ability to grow intracellularly than purine biosynthesis mutants reported previously. An EGDe (serotype 1/2a) mutant in purA exhibited a 3.4-fold (<1 log10) reduction in intracellular growth (Schauer et al., 2010), in comparison to the approximately 3 log10 reduction that we observed with ROA14 (purA mutant) and J22F (purB mutant). However, in our study the Caco-2 cells were incubated for 15–18 days, which results in polarized monolayers (Neudeck et al., 2004), whereas Schauer et al. (2010) used Caco-2 cell monolayers within 22 h after seeding. Intracellular growth of the mutants in the current study was also impaired to a greater extent than reported previously for an adenine auxotroph of strain 10403S in Henle 407 and J774 cells (Marquis et al., 1993). Such distinctions may reflect differences (e.g., serotype and lineage) in the strains employed in the studies and the cell types employed for evaluating invasion
More recently, there have been novel lines of evidence for the importance of purines in properties related to virulence of L. monocytogenes. For example, production of the signaling molecule cyclic-di-AMP by L. monocytogenes was found to result in production of interferon-β by infected macrophages (Woodward et al., 2010). This response is dependent on the signaling protein STING in macrophages (Sauer et al., 2011).
Our observation that purA and purB mutants of L. monocytogenes poorly colonize the gastrointestinal tract (i.e., cecum) are interesting in light of a recent report by Dowd et al. (2011) that purB is required for growth of L. monocytogenes EGDe in porcine gallbladder bile. However, we did not note a significant defect in the ability of the purB mutant to survive in bile-containing synthetic gastric fluid. A possible explanation is that synthetic gastric fluid contains bile salts, whereas Dowd et al. (2011) used raw bile harvested from pig gallbladders. Differences (including serotype) in the strains employed may also contribute to the differing results.
Conclusion
In summary, the results of this study provide evidence that purine biosynthesis is critical for full expression of virulence by L. monocytogenes serotype 4b in A/J mice inoculated via the gastrointestinal tract. Based on in vitro findings with Caco-2 cells, we infer that this may reflect in part a reduced ability of pur mutants to multiply within intestinal epithelial cells. Further studies are needed to elucidate the mechanisms mediating the critical role of purine biosynthesis in virulence and pathogenesis of serotype 4b L. monocytogenes.
Footnotes
Acknowledgments
We are grateful to H. Marquis (Cornell University) for the gift of pMC39. We thank R.M. Silertzky (the laboratory of S.K.) and all other members of our laboratories for their encouragement and support. This work was partially supported by the USDA National Alliance for Food Safety and Security (cooperative agreement number 59-1935-3-371), the National Research Initiative of the USDA Cooperative State Research, Education and Extension Service (grants 2005-35201-15313 to C.J.C. and 2006-35201-17377 to S.K.), the Walter and Martha Renk Endowed Laboratory for Food Safety (C.J.C.), and the UW-Madison Food Research Institute.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.