
Abstract
The objective of this research was to determine minimal inhibitory concentration (MIC) population distributions for colistin for Salmonella on subtype level. Furthermore, we wanted to determine if differences in MIC for colistin could be explained by mutations in pmrA or pmrB encoding proteins involved in processes that influence the binding of colistin to the cell membrane. During 2008–2011, 6,583 Salmonella enterica subsp. enterica isolates of human origin and 1931 isolates of animal/meat origin were collected. The isolates were serotyped, and susceptibility was tested towards colistin (range 1–16 mg/L). Moreover, 37 isolates were tested for mutations in pmrA and pmrB by polymerase chain reaction (PCR) and DNA sequencing. MIC distribution for colistin at serotype level showed that Salmonella Dublin (n=198) followed by Salmonella Enteritidis (n=1247) were less susceptible than “other” Salmonella serotypes originating from humans (n=5,274) and Salmonella Typhimurium of animal/meat origin (n=1794). MIC was ≤1 mg/L for 98.9% of “other” Salmonella serotypes originating from humans, 99.4% of Salmonella Typhimurium, 61.3% of Salmonella Enteritidis, and 12.1% of Salmonella Dublin isolates. Interestingly, Salmonella Dublin and Salmonella Enteritidis belong to the same O-group (O:1, 9,12), suggesting that surface lipopolysaccharides (LPS) of the cell (O-antigen) play a role in colistin susceptibility. The epidemiological cut-off value of >2 mg/L for colistin suggested by European Committee on Antimicrobial Susceptibility Testing (EUCAST) is placed inside the distribution for both Salmonella Dublin and Salmonella Enteritidis. All tested Salmonella Dublin isolates, regardless of MIC colistin value, had identical pmrA and pmrB sequences. Missense mutations were found only in pmrA in one Salmonella Reading and in pmrB in one Salmonella Concord isolate, both with MIC of ≤1 for colistin. In conclusion, our study indicates that missense mutations are not necessarily involved in increased MICs for colistin. Increased MICs for colistin seemed to be linked to specific serotypes (Salmonella Dublin and Salmonella Enteritidis). We recommend that Salmonella with MIC of >2 mg/L for colistin be evaluated on the serovar level.
Introduction
Previously, colistin was used less frequently due to toxicity, but recently it has become the first choice treatment of severe infections involving some multidrug resistant Gram-negative bacteria (Lim et al., 2010).
We hypothesized that colistin MIC population distributions of Salmonella on serotype level differ and MICs of colistin are increased in Salmonella isolates with missense mutations in pmrA or pmrB encoding products involved in processes that influence cell membrane binding of colistin (Roland et al., 1993; Sun et al., 2009).
The objective of this work was to determine MIC population distributions for Salmonella on subtype level and to determine if differences in MIC could be explained by mutations in pmrA or pmrB.
Methods
From 2008 to 2011, the MIC for colistin (concentration range 1–16 mg/L) was determined using commercially dehydrated antimicrobial agents in microtiter wells (Sensititre™; TREK Diagnostic Systems Ltd., West Sussex, United Kingdom) as part of the Danish Integrated Antimicrobial Resistance Monitoring and Research Programme (DANMAP) (CLSI, 2008; DANMAP, 2008–2010). From human patients in Denmark, 6,583 Salmonella enterica subsp. enterica isolates were tested. Among these isolates, the most common serotypes were Salmonella Typhimurium (3,281 isolates, including monophasic variants S. 4,5,12:i:- and S. 4,12:i:-), Salmonella Enteritidis (1,178 isolates), and Salmonella Dublin (131 isolates). The remaining 1,993 isolates belonged to 198 different serotypes. Furthermore, 1,795 Salmonella Typhimurium, 69 Salmonella Enteritidis, and 67 Salmonella Dublin from food animals and raw meat were tested.
In order to determine if differences in MIC were caused by mutations in pmrA and pmrB, we looked for mutations in 37 isolates (with MIC and identification verified) of serovar Salmonella Dublin (n=22), Salmonella Enteritidis (n=14), and Salmonella Typhimurium (n=1) from human infections in 2009 with MIC values of 2, 4, or 8 mg/L. In addition, one isolate of each of the above-mentioned three serotypes, plus one Salmonella Concord and one Salmonella Reading with MIC of ≤1 were included to look for differences in susceptible isolates of five different serotypes. The pmrA and pmrB genes were sequenced using the primers reported by Sun et al. (2009) and additional new primers designed to fully include pmrB (forward: 5′-CGACGGACTCAATCTCAAG-3′; reverse: 5′-GCATATACATAATTTGCGCGA-3′).
Results and Discussion
The MIC distribution for all Salmonella isolates is shown in Figure 1A; the MIC distributions for Salmonella Dublin, Salmonella Enteritidis, Salmonella Typhimurium (food animals and raw meats), and “other” serotypes originating from humans are presented in Figure 1B. Salmonella Dublin followed by Salmonella Enteritidis were less susceptible than the other Salmonella serotypes. Interestingly, Salmonella Dublin and Salmonella Enteritidis have the same O-antigen formula (O:1, 9,12).
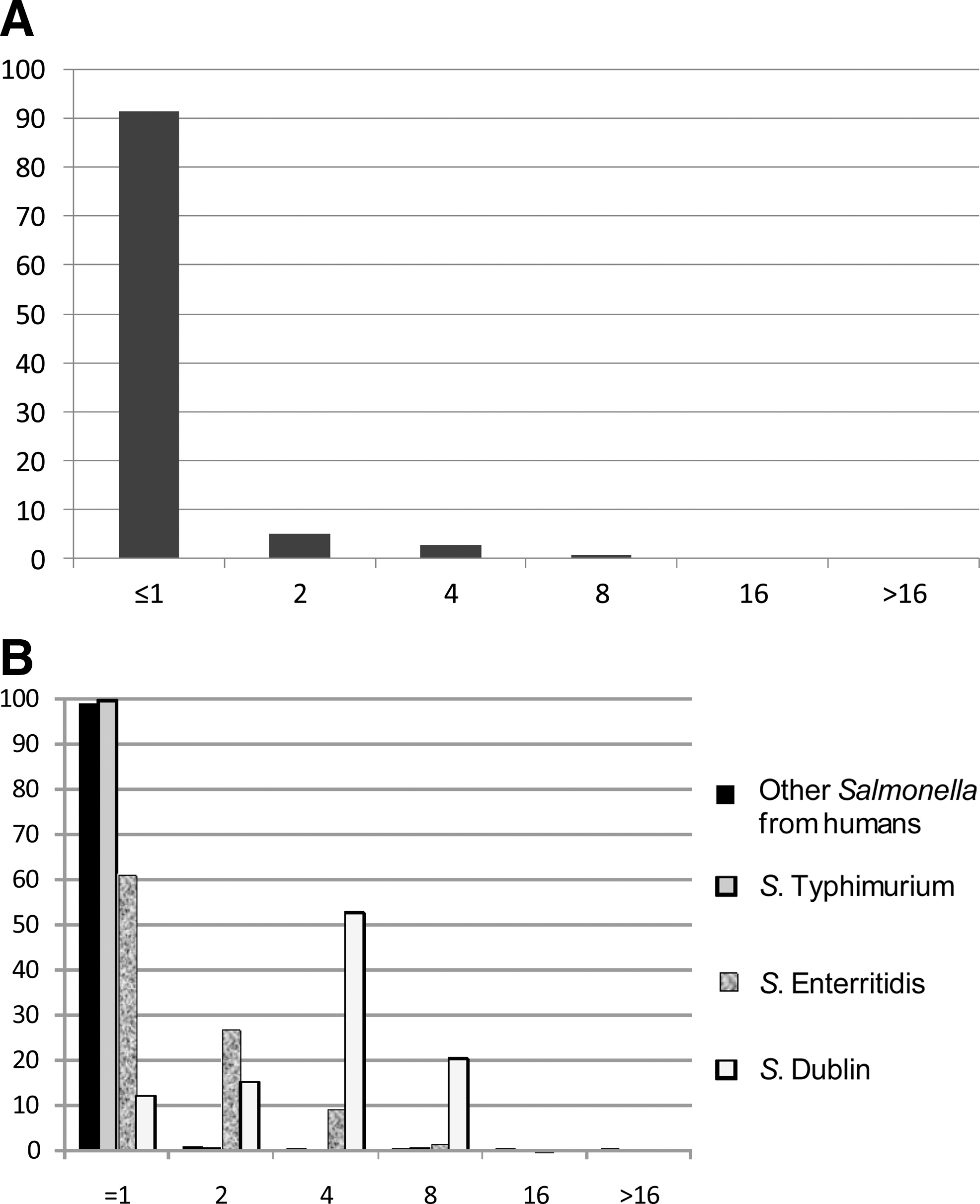
Therefore, we also looked at the MIC distribution of colistin for available Salmonella Napoli (n=18) and Salmonella Panama (n=30), which also have this O-antigen formula. Although these two serovars also seemed less susceptible to colistin than Salmonella in general, the trend was not as pronounced; MIC was ≤1 mg/L for 98.9% of “other” Salmonella serotypes of human origin, 99.4% for Salmonella Typhimurium from animal/meat origin, 89% for Salmonella Napoli, 80% for Salmonella Panama, 61.3% for Salmonella Enteritidis, and 12.1% for Salmonella Dublin. This suggests that the surface lipopolysaccharides (LPS) of the cell (O-antigen) play a role in colistin susceptibility. The epidemiological cut-off value suggested by EUCAST (>2 mg/L) is placed inside the distribution for both Salmonella Dublin and Salmonella Enteritidis. Due to the low number of especially Salmonella Dublin when compared to other Salmonella, these differences related to serotypes are overlooked when merging all Salmonella into the same distribution (Fig. 1).
Colistin targets the cell envelope, and the interaction between the cationic polypeptide and negatively charged LPS leads to a disturbance of the outer membrane and a subsequent increase in the permeability of the cell envelope resulting in death (Schindler and Osborn, 1979; Sun et al., 2009). Increase in MIC for Salmonella Typhimurium has been mapped to pmrA and pmrB, which constitute a two-component regulatory system. PmrB is a sensor histidine kinase, and PmrA is the cognate response regulator. The modifications in these genes influence the regulation of proteins, which makes the LPS less negatively charged and thereby decrease the binding of colistin (Roland et al., 1993; Sun et al., 2009).
Sun et al. (2009) found 27 missense mutations in pmrB and pmrA that lead to an increased MIC to colistin of up to 4.4 mg/L. For the isolates tested in the present study, all Salmonella Dublin isolates, regardless of the MIC value, had identical pmrA and pmrB sequences. Compared to the Salmonella LT2 strain (GenBank accession no. NC003197), the Salmonella Dublin isolates had a point mutation in pmrB position 25 (A→C), leading to an amino acid change in position 9 from threonine to proline. None of the other isolates had this mutation. In addition, five and six synonymous point mutations were seen in pmrB Salmonella Dublin and Salmonella Enteritidis isolates, respectively (two of them were identical, positions 492 and 750). No missense mutation was present in pmrA, but synonymous point mutations were seen at positions 177 and 322 in Salmonella Dublin (C→T) and at positions 322 and 414 in Salmonella Enteritidis (C→T). In addition, missense mutations in pmrA were found in one Salmonella Reading isolate, and a missense mutation in pmrB was found in one Salmonella Concord isolate; both had MIC of ≤1. Therefore, missense mutations are not necessarily involved in increased MICs for colistin.
In conclusion, since increased MIC seemed linked to serotype, we recommend that isolates with MIC of >2 mg/L for colistin in Salmonella spp. be evaluated on the serotype level. More detailed data when making population distributions (such as interpreting results based on the subtype level and looking for resistance mechanisms) may be necessary when determining interpretive criteria.
Footnotes
Acknowledgments
The technical staff at Statens Serum Institut and the National Food Institute, Technical University of Denmark are thanked for their assistance. We would like to thank Gunnar Kahlmeter for useful discussion. Data will be submitted to EUCAST for use in population distributions for Salmonella on serovar level. This work was supported in part by the Danish Ministry of Science, Technology and Innovation and the Danish Ministry of Health and Prevention as part of the Danish Integrated Antimicrobial Resistance Monitoring and Research Programme (DANMAP).
Disclosure Statement
No competing financial interests exist.