
Abstract
Shiga-toxigenic Escherichia coli O157:H7 outbreaks have been linked to consumption of fresh produce. It is generally recognized that bacterial attachment to vegetal matrices constitutes the first step in contamination of fresh produce. Cellular appendages, such as curli fibers, and cellulose, a constituent of extracellular matrix, have been suggested to be involved in E. coli attachment and persistence in fresh produce. A comparative evaluation was conducted on the ability of Shiga toxin-producing E. coli O157:H7 strains EDL933 and 86-24, linked to two independent foodborne disease outbreaks in humans, and their mutants deficient in curli and/or cellulose expression to colonize and to firmly attach to spinach leaf. Inoculated spinach leaves were incubated at 22°C, and at 0, 24, and 48 h after incubation loosely and strongly attached E. coli O157:H7 populations were determined. Curli-expressing E. coli O157:H7 strains developed stronger association with leaf surface, whereas curli-deficient mutants attached to spinach at significantly (p<0.01) lower numbers. Attachment of cellulose-impaired mutants to spinach leaves was not significantly different from that of curliated strains. The relative attachment strength of E. coli O157:H7 to spinach increased with incubation time for the curli-expressing strains. Laser scanning confocal microscopy (LSCM) analysis of inoculated leaves revealed that curli-expressing E. coli O157:H7 were surrounded by extracellular structures strongly immunostained with anti-curli antibodies
Introduction
T
Methods
Plant material
Organic spinach (Emilia cultivar) seeds were obtained from a commercial seed supplier (Seedway, Hall, NY). Approximately 300 seeds (2.84 g) were surface sterilized for 30 min with 1.0% sodium hypochlorite (5%; Sigma, St. Louis, MO), followed by five washes with sterile deionized water. To promote uniform and vigorous germination, seeds were primed in 200 mL of solution containing 30% (wt/vol) polyethylene glycol (PEG) 8000 (Sigma) for 72 h as recommended by Hart et al. (2006). After priming, PEG was removed from the seeds by washing, and seeds were planted in a “cone-tainer” (Stuewe & Sons, Inc., Corvallis, OR) filled with Miracle-Gro garden soil (Miracle-Gro, Marysville, OH) and containing a new glass fiber wad at the base (to prevent soil from passing out of the drainage holes). Germination and seedling growth were carried out in a growth chamber at 22°C and a relative humidity of 50–60% using a photoperiod consisting of 18 h of light (600 μmol m–2 s–1) and 6 h of darkness. Spinach plants were irrigated as needed to maintain soil moisture and plant turgor.
Bacterial strains and mutant construction
Shiga toxin-producing E. coli O157:H7 wild-type strains and their mutants (lacking the ability to produce curli or cellulose or both) used in the current study are listed in Table 1. E. coli O157:H7 strain EDL933 and its isogenic mutants have been described in a previous study (Saldana et al., 2009). The csgA and csgD mutants of E. coli O157:H7 strain 86-24 were constructed by using the bacteriophage λ-red-mediated recombination method as described previously (Datsenko and Wanner, 2000). Two successive transfers of cryo-preserved (-80°C) E. coli strains were made in Luria Bertani broth (LB; Neogen, Lansing, MI) at 37°C for 24 h. Streptomycin (100 μg/ml), kanamycin (50 μg/ml), ampicillin (100 μg/ml), and chloramphenicol (34 μg/ml) were added to LB as required for the growth of bacterial strains used in this study (Table 1).
curli+, curli-expressing strain; curli
Congo red and calcofluor staining
To identify the optimal temperature for curli production, overnight cultures of two E. coli O157:H7 strains and their mutants were streaked on LB agar plates devoid of salt and supplemented with Congo Red (40 μg/mL; Sigma-Aldrich, St. Louis, MO) and Coomassie brilliant blue (20 μg/mL; Sigma-Aldrich) (Römling et al., 2003), and incubated at 18°C, 20°C, 22°C, 28°C, and 37°C for 48 h. Bacteria-producing curli developed red colonies as a result of dye uptake. To detect cellulose production, bacteria were grown on LB plates without salt containing 200 mg/L calcofluor (Uhlich et al., 2006) and incubated at 18°C and 22°C for 48 h with cellulose-producing colonies fluorescing under ultraviolet (UV) light.
Inoculum preparation
Bacterial strains (Table 1) were grown in 10 mL of LB broth at 37°C and, when required, supplemented with antibiotics: kanamycin (50 μg/mL), streptomycin (100 μg/mL), chloramphenicol (30 μg/mL), and ampicillin (200 μg/mL). After 24 h of growth, 1 mL of bacterial culture was centrifuged at 5000×g for 15 min. Pelleted cells were washed with phosphate-buffered saline (PBS) and resuspended in 40 mL of PBS. Spinach leaves of approximately the same size (approximately 2×3 cm) were harvested from 5-week-old plants and then spot inoculated with 100 μL of suspension (5 spots of 20 μL of inoculums per leaf) to obtain 7 log colony forming units (CFU) of bacteria/leaf. Uninoculated leaves were used as controls.
Adherence assays
At 0, 24, and 48 h after incubation, loosely and strongly attached E. coli O157:H7 populations were determined as described by Patel et al. (2011). Briefly, inoculated spinach leaves were washed in sterile PBS to remove unattached bacteria; leaves were transferred to 50-mL conical tubes filled with 20 mL of sterile PBS containing 0.05% Tween 20 (PBS-TW) and vortexed for 2 min to dislodge loosely attached bacteria. After vortexing, leaves were transferred to new 50-mL conical tubes containing 20 mL PBS-TW and homogenized using a Polytron® 2100 homogenizer (Kinematica, Littau-Lucerne, Switzerland) to determine populations of strongly attached bacteria. PBS (loose attachment) and homogenate (strong attachment) were spiral plated on SMAC agar supplemented when required with a corresponding antibiotic (Table 1). After overnight incubation at 37°C, presumptive E. coli O157:H7 colonies were counted using automated colony counter (ProtoCOL; Microbiology International Inc., Frederick, MD). Randomly selected colonies were confirmed for E. coli O157 lipopolysaccharide (LPS) antigens by latex agglutination assay (Dryspot assay; Oxoid Ltd., Cambridge, UK). The relative attachment strength (SR) was determined as earlier described by Dickson and Koohmaraie (1989) and represents the ratio of the strongly attached bacterial cells to the total population of bacteria associated with the produce surface [SR=(strongly attached bacteria)/(strongly attached+loosely associated bacteria)].
LSCM analysis
At 24 and 48 hpi (hours post inoculation), leaves were washed three times in PBS to remove unattached bacteria and fixed in −40°C methanol for 5 min, followed by three washes with PBS and incubation for 30 min in PBS containing 2% non-fat dry milk to block non-specific immunoglobulin binding in subsequent steps. After blocking, the samples were incubated with rabbit anti-curli polyclonal antibodies (1:1000 dilution) (Saldana et al., 2009) for 2 h at 37°C, followed by 2-h incubation with fluorescein isothiocyanate (FITC)–conjugated goat-anti-rabbit immunoglobulin G (IgG, 1:50 dilution; Sigma). All antibodies were diluted in PBS-Tween 20 (PBS-TW), and removed after each step by three washes with PBS-TW. A Zeiss 710 LSCM system with 63× and 100×1.4 NA oil immersion Plan Apochromatic objectives was utilized in immunofluorescence analysis. Differential interference contrast (DIC) and confocal fluorescence images were acquired simultaneously. A photomultiplier tube captured the light emitted from a 488-nm argon laser with a 3.7-μm pin hole passing through an MBS 488 filter with limits set between 492 and 543 nm for detection of fluorescein and between 647 and 721 nm for detection of autofluorescence from chloroplasts. Zeiss Zen 2008 software was used to obtain the images with 512×512 pixel resolution.
Statistical analysis
A randomized complete block design was used with three replicates per treatment. The populations of loosely and strongly attached E. coli O157:H7 obtained at each sampling period were converted to log CFU/g. The data obtained from three replicates were analyzed by a two-way analysis of variance (ANOVA) using Proc Mixed (SAS 8.2; SAS, Cary, NC) for interaction effects of the strain and sampling period. The results were considered statistically significant at p<0.01.
Results
The highest levels of curli production were observed after incubation at 22°C. E. coli O157:H7 strains EDL933ΔcsgA, EDL933ΔcsgD, EDL933ΔcsgA/ΔbcsA, and 86-24ΔcsgA mutants deficient in curli or curli and cellulose production did not develop red phenotype on Congo red media at any incubation temperature used in the study. Among the curli-producing and Congo red-binding strains, the strongest level of curli expression was observed for the wild-type E. coli O157:H7 strain 86-24 and its constitutive curli mutant 86-24 csgDc (Fig. 1). All strains, except EDL933ΔbcsA and EDL933ΔbcsA/ΔcsgA, produced cellulose (data not shown). Since the greatest level of curli expression was observed at 22°C (also accompanied by cellulose biosynthesis), this temperature was used in further analysis of bacterial attachment to spinach leaves.

Phenotypic presentation of Escherichia coli O157:H7 curli variants grown on Luria Bertani broth (LB; Neogen, Lansing, MI)/no-salt medium supplemented with Congo Red at 22°C for 48 h. The intensity of colony pigmentation as a result of Congo Red binding indicates different level of curli production. Color images available online at
Immediately after inoculation (Time 0), numbers of loosely attached bacteria were not significantly different among the E. coli O157:H7 strains, ranging from 4.2 to 4.5 log CFU/leaf (data not shown). After 24-h incubation, numbers of loosely attached bacteria were similar to those observed at time 0, with the exception of EDL933ΔcsgD, which showed significantly (p>0.01) lower recovery (3.7 log CFU/leaf). After 48-h incubation, numbers of loosely attached bacteria on the spinach leaves were in the range of 4.5 to 4.9 log CFU/ leaf and did not vary significantly (p>0.01) among mutants and parental strains. Recovery of loosely attached bacteria indicated that, after undergoing initial die-off, overall curli- and cellulose-impaired strains maintained relatively stable populations of loosely attached cells similar to those of curli- and cellulose-expressing genotypes; nevertheless curli-negative EDL933csgD mutant (deficient in CsgD transcriptional activator) showed the least attachment to leaf surface at 24 h post inoculation.
At time 0, populations of strongly attached bacteria recovered from the spinach leaves ranged from 2.2 to 3.3 log CFU/leaf (0 hpi; Fig. 2). Curli-expressing E. coli O157:H7 strains developed stronger association with leaf surface with time, as indicated by the presence of significantly higher numbers of strongly attached cells (4.1–4.8 log CFU/leaf) by 24 h of incubation (Fig. 2). Curli-deficient mutants, however, failed to attach at higher numbers after 24 h. Strongly attached populations of E. coli O157:H7 strains EDL933ΔcsgA (3.2 log CFU/g) and EDL933ΔcsgA/ΔbcsA (3.6 log CFU/g) were similar to their corresponding populations at time 0. Populations of strongly attached curli-impaired mutants EDL933ΔcsgD and 86-24ΔcsgA decreased within 24 h to 3.0 and 2.4 log CFU/leaf, respectively. After 48 h, numbers of curli-deficient bacteria strongly attaching to spinach have increased.
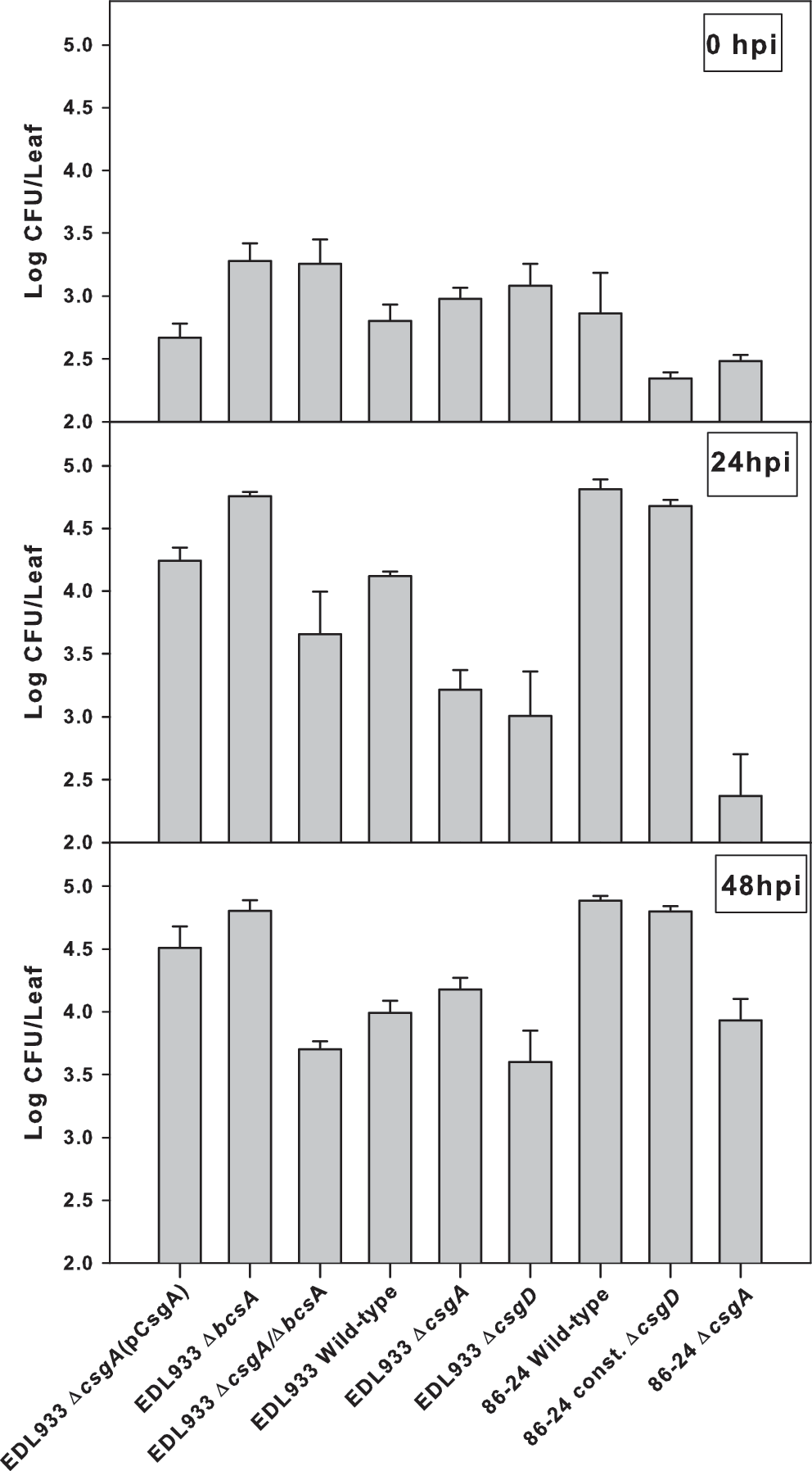
Populations of strongly attached Escherichia coli O157:H7 and its curli and cellulose mutants to spinach leaf incubated at 22°C. Counts (Log colony forming unit [CFU]/leaf) obtained from Sorbitol MacConkey (SMAC) agar containing antibiotics (as needed). Error bars are the standard deviations from three replications. hpi, hours post inoculation.
At 0 hpi, the attachment strength (SR) of E. coli O157:H7 to spinach was of low and variable magnitude (0 hpi in Fig. 3). With time, attachment strength significantly increased for the curli-expressing strains; at 24 h, the difference in SR values between curli-producing and curli-deficient strains ranged from 1.5 to 200-fold (24 hpi in Fig. 3). At 48 h, the differences in the attachment strengths remained significant among curli-competent and curli-impaired E. coli O157:H7 strains (48 hpi in Fig. 3).
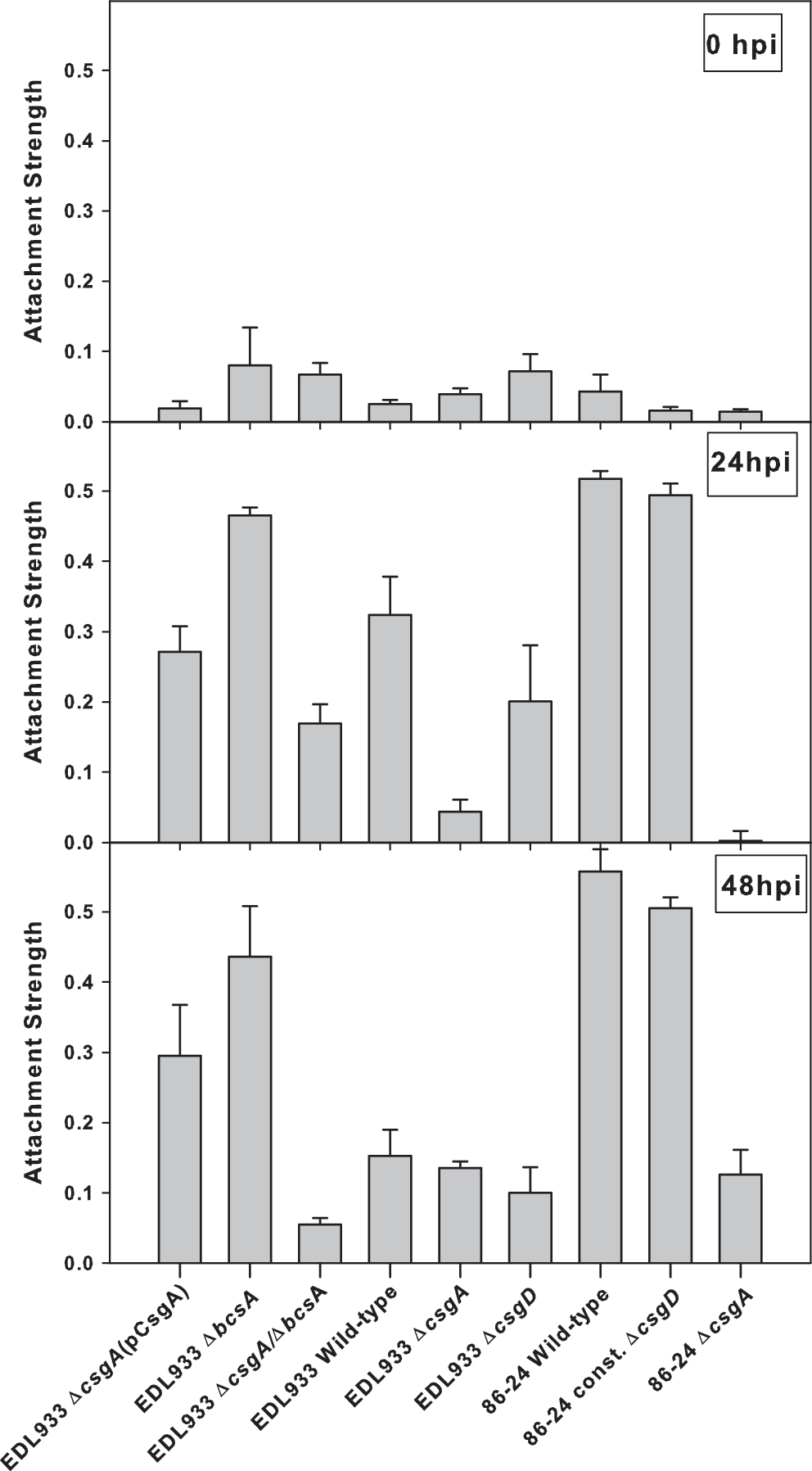
Attachment strength of Escherichia coli O157:H7 and its curli and cellulose mutants to spinach leaf incubated at 22°C. The relative attachment strength (SR) values were calculated as the ratio of strongly attached cells/(strongly+loosely attached cells). Error bars indicate the standard deviations from three replications. hpi, hours post inoculation.
LSCM analysis of inoculated spinach leaves revealed that bacteria firmly attached on leaf surface were surrounded by an extracellular matrix as recognized by rabbit anti-curli antibodies. Figure 4 shows leaf surface colonized by E. coli O157:H7 constituitive mutant 86-24csgD c (Fig. 4A–C) and wild-type parental strain 86-24 (Fig. 4D–J), both intensely immunostained for curli. Curli-imparied EDL933ΔcsgD and 86-24ΔcsgA mutant strains, which attached at a significantly lower numbers (p<0.01) on spinach at 24 hpi, were not recognized by anti-curli polyclonal antibodies (Fig. 5). Immunoflorescence analysis of inoculated spinach leaves did not detect curli expression by EDL933ΔcsgA/ΔbcsA, EDL933ΔcsgA, EDL933ΔcsgD, or 86-24ΔcsgA, at 48 hpi (data not shown).
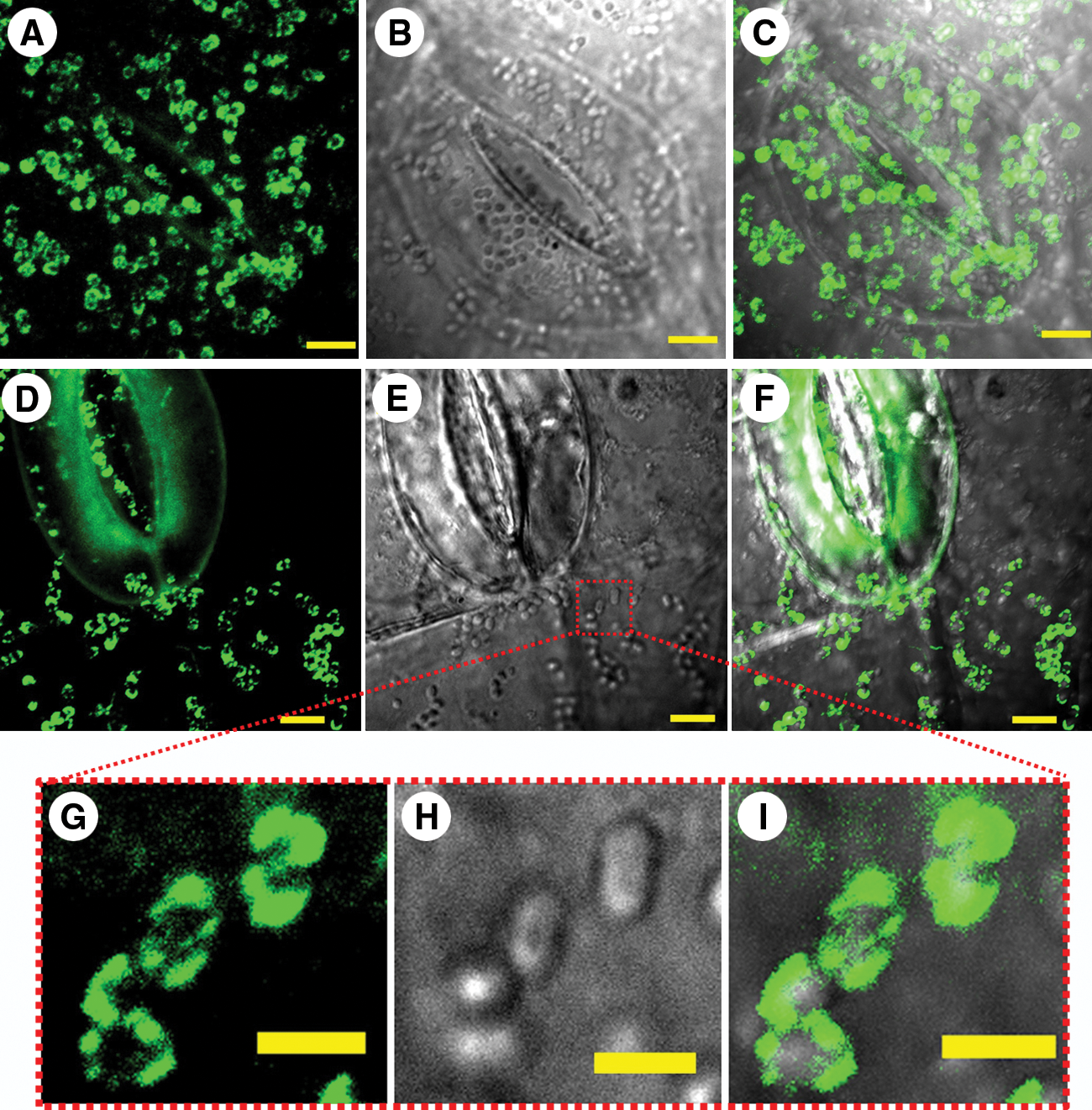
Laser scanning confocal microscopy (LSCM) images of a constitutive curli mutant 86-24 csgDc
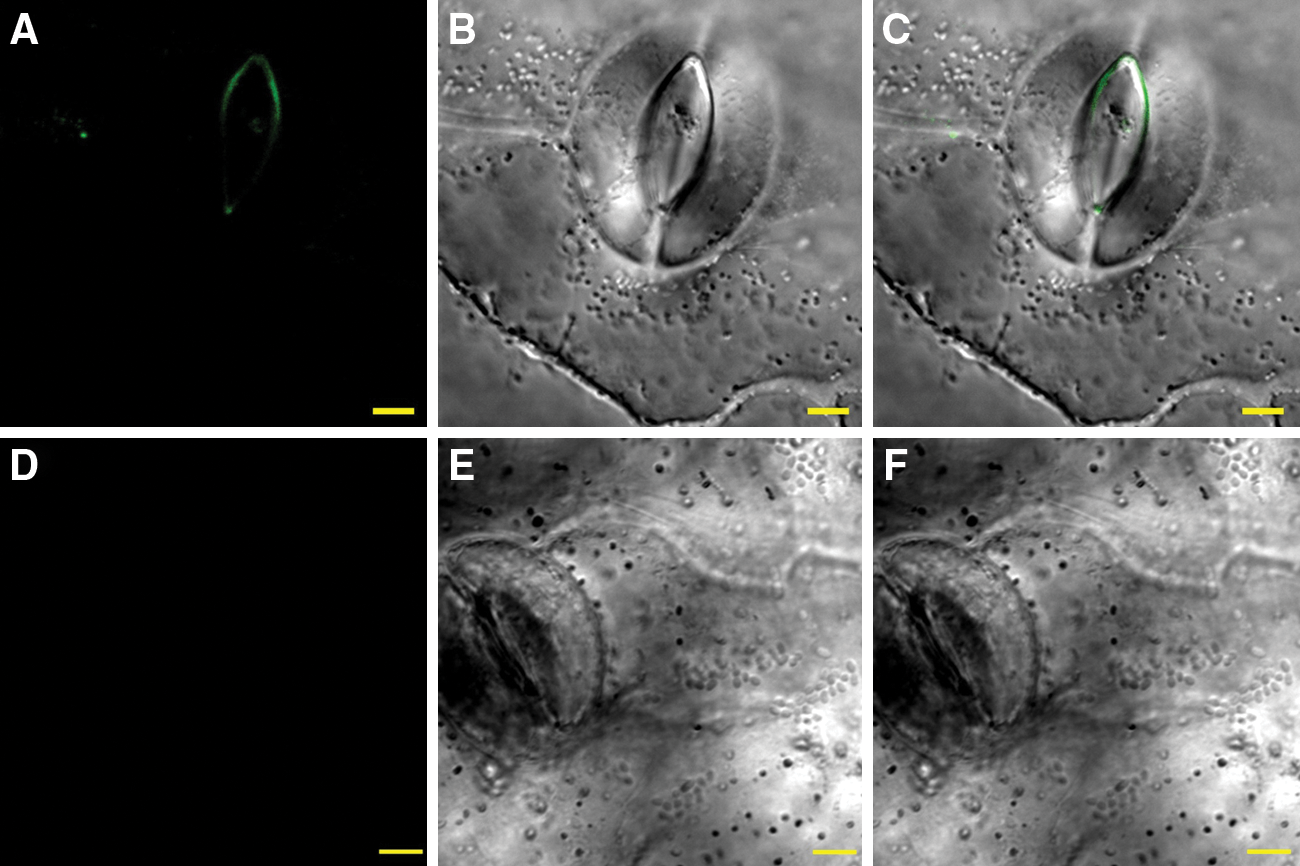
Laser scanning confocal microscopy (LSCM) images of Escherichia coli O157:H7 curli-deficient mutants EDL933ΔcsgD
Discussion
The present study was conducted to address the potential use of curli and cellulose by E. coli O157:H7, an important foodborne human pathogen, to colonize and persist on fresh produce. Specifically, the effect of curli and cellulose expression on E. coli O157:H7 attachment to spinach leaves was evaluated. Earlier research has supported the involvement of these cellular appendices in E. coli O157:H7 attachment to leafy greens (Torres et al., 2005; Jeter and Matthysse, 2005; Matthysse et al., 2008; Patel et al., 2010; Saldana et al., 2011). On the other hand, there are studies showing no role for curli and cellulose in colonization and persistence on plant surfaces (Uhlich et al., 2006; Boyer et al., 2007). Production of curli and cellulose varied among E. coli strains and was highly responsive to external stimuli (Carter et al., 2011) which could have explained differences in results of these studies. To minimize the strain effects in evaluating the role of curli and cellulose expression in E. coli O157:H7 attachment to spinach leaves, two lineages of well-studied enterohemorrhagic E. coli O157:H7 strains, EDL933 and 86-24, and their isogenic mutants were included in the current study. Also, to reduce the number of variables while assessing the role of cellular appendices in bacterial colonization of leaf surface, inoculated spinach was incubated at 22°C, an optimal temperature for curli and cellulose production by these strains. Further studies are needed to determine the role of curli and cellulose in bacterial adherence to and biofilm formation on produce surfaces at 4°C in commercial storage settings.
Experiments on the recovery of loosely attached bacteria to spinach leaves showed that the populations of curli- and cellulose-positive strains were comparable to those of curli- and cellulose-negative mutants at all sampling periods, suggesting that production of curli and cellulose or lack thereof by strains EDL933 and 86-24 had no effect on loose attachment of E. coli O157:H7 to spinach. Recovery of strongly attached curli-deficient mutants, EDL933ΔcsgA/ΔbcsA, EDL933ΔcsgA, EDL933ΔcsgD, and 86-24ΔcsgA, were significantly lower than the recovery of strongly attached curli-expressing mutants and wild-type parental strains. In a recent study, Saldana et al. (2011) reported that, among known adherence factors (such as E. coli common pilus, T3SS, flagella), E. coli O157:H7 strain EDL933 also utilizes curli to colonize baby spinach leaves. In our study, curli were critical in strong attachment to spinach, whereas loose attachment to spinach was not affected by curli expression. The differences could be due to the different methodology used in quantitative assessment of bacterial attachment. Unlike the work of Saldana et al. (2011), this study differentiated bacterial attachment to spinach leaves in loose and strong attachment.
In our study, lack of cellulose expression by EDL933ΔbcsA did not affect its ability to develop firm binding to leaf surface. Integral to that, deletion of the bcsA gene in a curli/cellulose-impaired mutant (EDL933ΔcsgA/ΔbcsA) did not reduce its binding capacity to spinach as compared to non-curliated EDL933ΔcsgD and 86-24ΔcsgA. Our results are in agreement with Uhlich et al. (2006), who reported that cellulose biosynthesis was not necessary in E. coli O157:H7 attachment and biofilm formation on inanimate surfaces.
Attachment strength (SR) correlated with bacterial ability to produce curli. Both wild-type strains and their curliated mutants had significantly higher SR values as compared to curli-deficient strains. Notably, the strength of attachment of E. coli O157:H7 to Romaine lettuce surface also positively correlated with curli expression (Patel et al., 2011). The differences in the attachment strength between curli negative and curli-producing strains increased with incubation time, which were likely due to the gradual increases in curli biosynthesis in the curli-positive strains resulting in stronger adherence to leaf surfaces. Our data are in agreement with previous study showing a significant increase in the attachment strength of curli-expressing Salmonella serovars to produce surfaces within a 24-h time interval (Patel et al., 2010). The attachment strength of cellulose-deficient mutant strain EDL933ΔbcsA was not significantly different from the attachment strength of cellulose-expressing strains, indicating that this polysaccharide is not critical for E. coli O157:H7 attachment to spinach. This observation disagrees with an earlier report showing role of cellulose in bacterial colonization of plant surfaces (Barak et al., 2007). Differences in current results and those from the Barak et al. (2007) study may be due to different mechanisms of attachment between Salmonella and E. coli O157:H7.
In conclusion, the present study shows that loose attachment of E. coli O157:H7 to spinach leaves is not affected by extracellular appendices such as curli and cellulose. However, strong attachment of E. coli to plant surface during the first 24 h after inoculation is greatly influenced by its ability to produce curli fimbriae. A sharp increase in populations of the curli-expressing E. coli O157:H7 within fist 24 h after inoculation, together with LSCM observation of curli-composed extracellular matrix on leaf surfaces, clearly indicate that curli are critical in strong attachment to fresh produce. Cellulose production is not required for strong attachment to the spinach leaf by E. coli O157:H7 strains used in this study. A moderate increase in populations of strongly attached cells of both curli-positive and curli-deficient E. coli strains after 48 h suggests that, in addition to curli, some other bacterial adherence factors (e.g., flagella [Xicohtencatl-Cortes et al., 2009] or the type 3 secretion system [Saldana et al., 2011]) become involved at subsequent stages in persistent association with the spinach leaf.
Disclosure Statement
No competing financial interests exist.