
Abstract
The aims of this study were to determine whether Shigella species, which are human gastrointestinal pathogens, can adhere to cattle recto-anal junction squamous epithelial (RSE) cells using a recently standardized in vitro adherence assay, and to compare their adherence patterns with that of Escherichia coli O157. Shigella dysenteriae (serogroup A), S. flexneri (serogroup B), S. boydii (serogroup C), and S. sonnei (serogroup D) were tested in adherence assays using both RSE and HEp-2 cells, in the presence or absence of D+mannose. Escherichia coli O157, which adheres to RSE cells in a Type I fimbriae-independent manner, was used as a positive control. Shigella serogroups A, B, D, but not C adhered to RSE cells with distinct adherence patterns in the presence of D+mannose. No such distinction could be made between the four Shigella serogroups based on the HEp-2 cell adherence patterns. Thus, this study provides evidence that certain Shigella serogroups adhere to RSE cells in a manner that is similar to the adherence pattern of E. coli O157. These unexpected observations of in vitro binding of these foodborne human pathogens to cells of the bovine gastrointestinal tract warrant evaluation of Shigella carriage by cattle using both experimental and observational studies, especially for serogroups B and D. Such studies are currently underway.
Introduction
Shigella serogroups are primarily transmitted via the fecal-oral route from person to person and are responsible for several foodborne/waterborne outbreaks, especially in crowded gatherings and day care centers (Hale et al., 1996; Niyogi, 2005). A seasonal pattern is associated with these outbreaks, with a majority of them occurring summer through fall, much like E. coli O157 outbreaks (CDC, 2008; Tauxe et al., 1995; WHO, 2009; Kosek et al., 2010). Although no animal reservoirs are currently associated with Shigella, the fact that, S. flexneri and S. sonnei were isolated from mesenteric lymph nodes of cattle at slaughter and bone marrow of a calf with septicemia, respectively, raises the possibility that ruminants such as cattle could be carriers of these human pathogens as well (Deom et al., 1954; Ueda et al., 1963). This supposition is further supported by the genetic similarity of Shigella to E. coli, including E. coli O157, and the establishment of cattle as reservoirs for other non-E. coli enteric pathogens such as Salmonella and Campylobacter species (Besser et al., 2005; Callaway et al., 2011).
In this study, we evaluated the ability of the four Shigella serogroups to adhere bovine gastrointestinal tract-derived cells in vitro. In particular, we employed an adherence assay that utilizes bovine recto-anal junction squamous epithelial (RSE) cells that was recently standardized in our laboratory to dissect patterns of adherence of E. coli O157 to the bovine gastrointestinal tract (Kudva et al., 2011).
Methods
Bacterial strains and animals
The four serogroups of Shigella evaluated in this study included Shigella dysenteriae (serogroup A; ATCC 11456A [American Type Culture Collection, Manassas, VA]), S. flexneri (serogroup B), S. boydii (serogroup C; ATCC 12027), and S. sonnei (serogroup D). In addition, a total of five Shigella serogroup B (ID# 01-002, 01-003, 01-007, 02-050, and 99060161) and two Shigella serogroup D (ID# 01-052, 08-001) clinical strains were tested to verify consistency of adherence patterns for those serogroups. All Shigella serogroup B and D strains were obtained from Dr. Phillip I. Tarr (Pediatric Gastroenterology, Washington University School of Medicine, St. Louis, MO). Escherichia coli O157 (O157) strain 86-24 (Shiga toxin [Stx] 1-negative, Stx 2-positive; motile; clinical isolate; National Animal Disease Center, Ames, IA) that binds RSE epithelial cells with a diffuse adherence pattern (Kudva et al., 2011) was used as the positive control. Escherichia coli strain K12 (ATCC 700926), a non-adherent strain, was the negative control.
HEp-2 culture and adherence assay conditions
HEp-2 cells (ATCC CCL-23) were cultured in RPMI 1640 (Invitrogen, Carlsbad, CA) medium supplemented with 10% fetal bovine serum (FBS; Thermo Scientific HyClone, Logan, UT), 50 μg/mL gentamicin (Thermo Scientific Pierce Protein, Logan, UT), and 2.5 mg/L amphotericin B (Sigma-Aldrich, St. Louis, MO), referred to as RPMI 1640+, at 37°C, 5% CO2, and 95% humidity until fully confluent. Subsequently, cells were trypsinized and aliquots used to determine cell counts, to seed flasks and chamber-slides, and to prepare freezer stocks as previously described (Freshney et al., 2000). Trypsinized HEp-2 cells were resuspended in RPMI 1640+ to a final concentration of 105 cells/mL, and 400 μL of the suspension was used to seed each well of the eight-well Permanox (Lab-Tek Chamber Slide System; Nunc, Rochester, NY) slides (one bacterial strain per slide). The slides were seeded and incubated 48 h (37°C, 5% CO2, and 95% humidity) to achieve 60–80% confluence. Meanwhile, bacteria (Shigella strains, E. coli O157, or E. coli K12) were grown overnight at 37°C without aeration in Dulbecco's Modified Eagle Medium (DMEM; Invitrogen) with low glucose (DMEM-LG) and plated on LB plates to determine counts in colony-forming units per milliliter (CFU/mL). The bacterial cells were then pelleted (5311×g, 4°C, for 20 min) and re-suspended in DMEM-no glucose (DMEM-NG) with 2.5% D+mannose, which was included to block Type 1 fimbriae mediated adherence (Ofek et al., 1978; Schaeffer et al., 1980; Rodrigues et al., 2009). In accordance with the bacterial concentration, the bacterial suspension was added to HEp-2 cells on the slides (one bacterial strain per slide) to give a final bacteria/cell ratio of 10:1 per well (e.g., 105 bacteria/104cells). Prior to addition of bacteria, RPMI 1640+ was aspirated from each well of the HEp-2 slide and the cells washed three times with 400 μL Dulbecco's phosphate-buffered saline (DPBS; Sigma). After additions of bacteria in DMEM-NG with 2.5% D+mannose, the slides were incubated at 37°C for 4 h. Every 2 h during the incubation period, the medium was aspirated and wells washed four times with DPBS before adding 400 μL of fresh DMEM-NG with 2.5% D+mannose and re-incubating. This ensured that only tightly adherent bacteria remained attached to the HEp-2 cells. After a total of 4 h of incubation and the final washes, the slides were fixed in 95% ethanol for 10 min at room temperature and stained as described below. The assay was done in triplicate for each isolate (24 wells in total per isolate), and HEp-2 slides with no added bacteria were processed likewise and used as controls to confirm absence of pre-existing bacteria. The assay was also conducted in the absence of D+mannose in order to compare results.
Isolation of RSE cells and adherence assay conditions
RSE cells were either isolated from recto-anal junction mucosal swabs (RAMS) or from recto-anal junction (RAJ) tissue taken at necropsy using a protocol standardized in our laboratory (Kudva et al., 2011). Briefly, 6–8 RAMS per animal were transported to the laboratory in 10 mL of DMEM-NG supplemented with 2.5% FBS, 100 μg/mL streptomycin– 100 U/mL penicillin (Pen-Strep; Invitrogen), and 50 μg/mL gentamicin (Invitrogen), referred to as DMEM(+)NG, on ice. Upon receipt, the swabs were vortexed, aseptically removed, and samples from one animal pooled together. Pooled samples were then filtered through a sterile 70-μm nylon cell strainer (BD Biosciences, Chicago, IL) into a sterile tube and centrifuged at 850×g at 4°C for 15 min to collect the RSE cells pellet with some fecal debris. The process of filtration and centrifugation was repeated twice to remove fecal debris, and the residual pellet was then resuspended in DMEM(+)NG. Cell counts and cell viability were determined as before using 0.4% Trypan blue staining (Freshney et al., 2000), and −80°C stocks of RSE cells were prepared using 50% glycerol (Sigma). In case of necropsies, the RAJ tissue samples were transported in DMEM(+)NG on ice, scraped using a slide to collect the surface epithelial cells in the laboratory, and the cells processed in the same manner as described for RAMS samples to harvest RSE cells.
Freshly isolated or frozen stocks of RSE cells, from multiple cattle, were pooled to rule out any age/breed bias and used in this assay, which was conducted in 16×100 mm glass tubes. RSE cells were washed in 14 mL of DMEM-NG with 2.5% D+mannose and filtered through a 70-μm nylon filter, twice, before resuspending them in 1 mL of the same media to a final concentration of 105 cells/mL. Bacteria (Shigella strains, E. coli O157, or E. coli K12) from overnight cultures in DMEM-LG, incubated at 37°C without aeration, were plated on LB plates to determine counts in CFU/mL. Subsequently, the bacteria were pelleted, resuspended in DMEM-NG with 2.5% D+mannose, added to the RSE cells suspension to a final bacteria/cell ratio of 10:1 (e.g., 106 bacteria/105 cells), and the mixture incubated with aeration (37°C, 110 rpm, for 4 h). At the end of 4 h, a 100-μL aliquot of the mixture was set aside to evaluate bacterial growth, in case no bacteria were to be observed on the slides post-staining (data not shown). Next, the mixture was pelleted and washed thoroughly, once with 14 mL of DMEM-NG and twice with 14 mL of sterile, distilled water (dH2O) before reconstituting in 100 μL of dH2O. Eight 2-μL drops of this suspension were spotted on Polysine (Thermo Scientific Pierce) slides and dried overnight under direct light to quench non-specific fluorescence (Neumann et al., 2002), before fixing in cold 95% ethanol for 10 min and staining. The assay was done in triplicate for each bacterial isolate (24 spots in total per isolate), and RSE cells with no added bacteria were subjected to the assay procedure and used as controls to confirm absence of pre-existing bacteria. The assay was also conducted in the absence of D+mannose in order to compare results.
Immunofluorescent (IF) staining
Ethanol-fixed slides were washed twice in phosphate buffered saline (PBS) at room temperature and blocked with 5% normal goat serum (Vector Laboratories, Burlingame, CA) in PBS (37°C for 30 min) prior to incubation with primary antibodies (37°C for 1 h). Primary antibodies, diluted in PBS, were combined as follows: mouse anti-(PAN) cytokeratins (AbD Serotec, Raleigh, NC) that target the cytokeratins of both the HEp-2 and RSE cells, with rabbit anti-Shigella (Kirkegaard Perry Laboratories, Gaithersburg, MD) that target the Shigella strains, or with rabbit anti-E. coli (Thermo Scientific Pierce) that target E. coli K12. Subsequently, slides were washed three times with PBS at room temperature and incubated with secondary antibodies diluted in 5% normal goat serum in PBS (37°C for 1 h).
Alexa Fluor 594 (red) labeled goat anti-mouse IgG (H+L; F(ab′)2 fragment; Invitrogen) targeting the anti-cytokeratins primary antibody was common to all secondary antibody combinations used. The other antibodies added along with this antibody included either Alexa Fluor 488 (green) labeled goat anti-rabbit antibodies (Invitrogen) targeting the anti-Shigella primary antibody, or the flourescein isothiocyanate (FITC; green)-labeled goat anti-O157 (KPL, Gaithersburg, MD) antibodies that labels O157. Following two washes with PBS and two washes with distilled water, the slides were air dried in the dark and coverslipped with Prolong Gold antifade reagent containing the DNA stain 4’,6’-diamidino-2-phenylindole (DAPI; Invitrogen). All slides were analyzed using the Nikon Eclipse E800 fluorescence microscope (Nikon Instruments, Elgin, IL) equipped with fluorescence illumination and digital imaging. Digital images were obtained using a CCD camera (Diagnostics Instruments, Sterling Heights, MI) and acquired using the MetaVue software (Molecular Devices, Sunnyvale, CA) as described previously (Kudva et al., 2011).
Control slides with HEp-2 or RSE cells, but no bacteria, were stained with the same sets of antibodies to rule out non-specific binding. Additional controls were prepared by staining test slides with unrelated fluorescence-tagged antibodies to demonstrate specificity of related antibody reactions. For the latter, slides were blocked with 5% normal rabbit serum (NRS; Vector Laboratories) and 5% normal horse serum (NHS; Vector Laboratories), and different combinations of primary and secondary antibodies were used. The mouse anti-(PAN) cytokeratins (AbD Serotec) and goat anti-Salmonella (Thermo Scientific Pierce) that targets all Salmonella strains comprised the primary antibody combination in PBS. Alexa Fluor 594 (red) labeled donkey anti-mouse IgG (H+L) (diluted in NHS; Invitrogen) targeting the anti-cytokeratins primary antibody was then combined with Alexa Fluor 488 (green) labeled rabbit anti-goat antibodies (diluted in NRS; Invitrogen) targeting the anti-Salmonella primary antibody to make the secondary antibody combination.
In addition, some of the HEp-2 adherence assay slides were stained with Alexa Fluor 594 (red) labeled phalloidin (Molecular Probes) after permeabilizing the cells with 0.1% Triton X in PBS, in place of anti-cytokeratin antibodies, in order to target the actin filaments in the cell (Knutton et al., 1989). This was done to verify any actin rearrangement associated with the invasive phenotype of Shigella species (Ally et al., 2004).
Qualitative and quantitative analysis of adherence
The HEp-2 slides were used for qualitative comparison of adherence patterns observed with RSE cells. Adherence patterns of all bacteria on HEp-2 and RSE cells were recorded as diffuse or aggregative (“stacked brick” appearance) for all positive interactions that involved direct association with the cells. Scattered bacteria and bacterial micro-colonies not adhering to cell membranes were considered to be negative for adherence to the epithelial cells (Doyle et al., 1995).
A total of 100–160 evenly dispersed RSE cells (10–20 RSE cells per spot) were analyzed per slide as previously described (Kudva et al., 2011). Percent RSE cells with and without bacteria adhering to them were determined. If 50–100% RSE cells had >10 bacteria attached to each cell, the adherence was recorded as strongly positive. For 50–100% RSE cells with 1–10 adherent bacteria per cell, the adherence was recorded as moderately positive. For less than 50% of RSE cells with 1–5 adherent bacteria per cell, the adherence was recorded as non-adherent.
Results and Discussion
Shigella species are foodborne pathogens that are transmitted via the feco-oral route through contaminated food and water (Hale et al., 1996; Niyogi, 2005). So far, houseflies have been implicated as transient carriers, but rarely have animals been identified as a reservoir for these human pathogens (Deom et al., 1954; Ueda et al., 1963; Hale et al., 1996). Given that cattle have been implicated as reservoirs of diverse enteric pathogens with human disease causing potential such as E. coli O157 and also given the genetic relatedness of Shigella to E. coli, (CDC, 2008; Tauxe et al., 1995; WHO, 2009; Hale et al., 1996), we sought to determine whether Shigella could specifically bind cells of bovine origin in vitro as a possible reflection of their ability to bind these cells in vivo. We therefore evaluated the adherence potential of the four Shigella serogroups using a bovine RSE cell adherence assay, recently standardized in our laboratory (Kudva et al., 2011).
Shiga toxin-producing E. coli (STEC) have been shown to colonize cattle at the RAJ among other sites and have been used to demonstrate adherence to explants/cells from this site (Naylor et al., 2003; Pohlenz et al., 2004; Baines et al., 2008; Kudva et al., 2011). Considering that Shigella is closely related to E. coli, we hypothesized that Shigella may show similar binding to these cells. Interestingly, Shigella serogroups A, B, and D did adhere to the RSE cells, with Shigella serogroup D exhibiting a distinct aggregative adherence pattern in the presence of D+mannose.
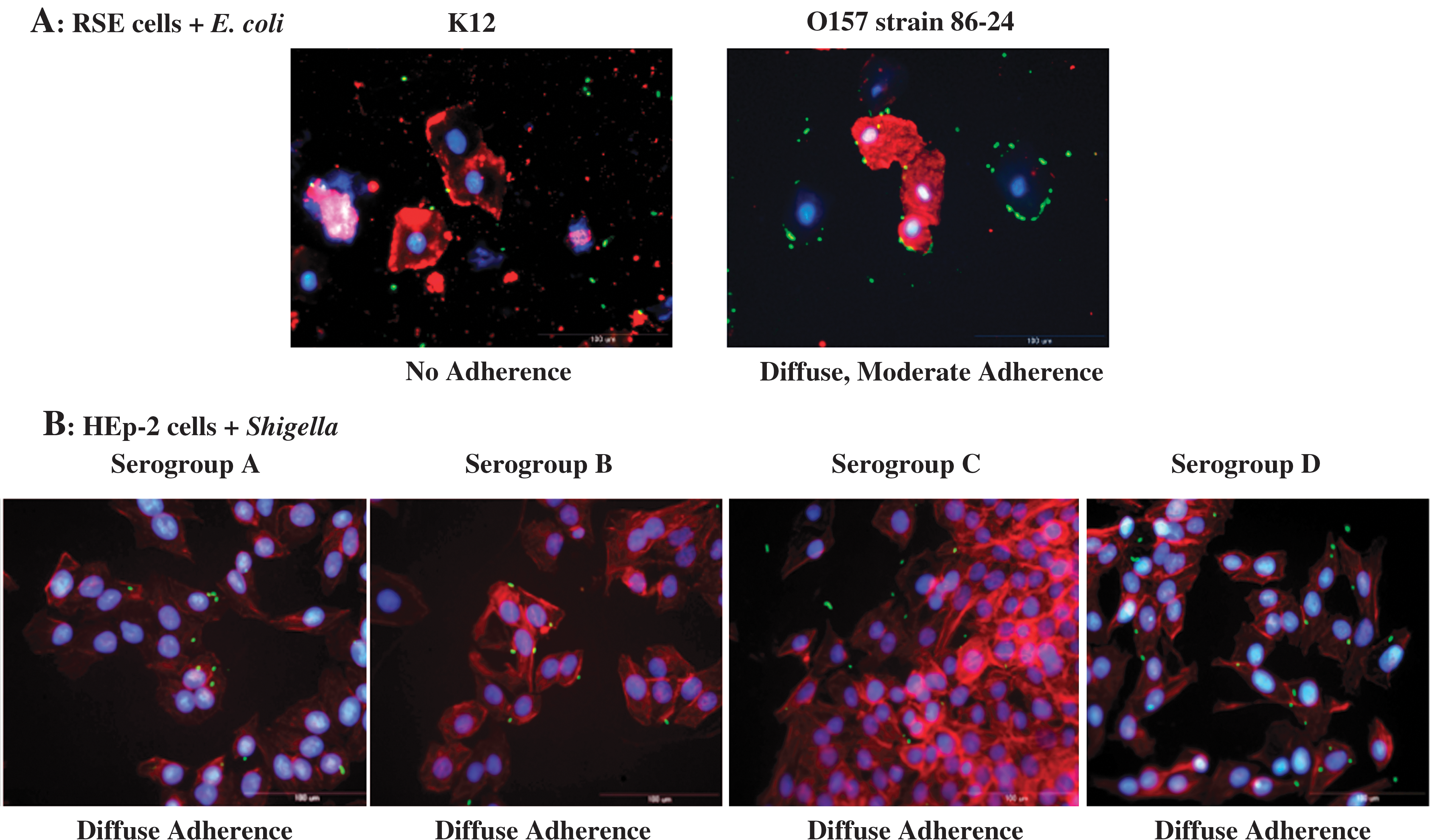
The RSE cell adherence assay was conducted in the absence or presence of D+mannose. In the absence of D+mannose, Shigella serogroups A, B, and C were moderately adherent, with bacteria binding 50–100% RSE cells in a diffuse adherence pattern, ranging from 1 to 10 bacteria/RSE cell (Fig. 1A). These adherence patterns of Shigella serogroups A, B, and C were similar to that observed with E. coli O157 strain 86-24 (Fig. 2A) (Kudva et al., 2011). On the other hand, Shigella serogroup D was strongly adherent, binding 50–100% RSE cells with a distinctive aggregative adherence pattern of >10 bacteria/RSE cell (Fig. 1A).
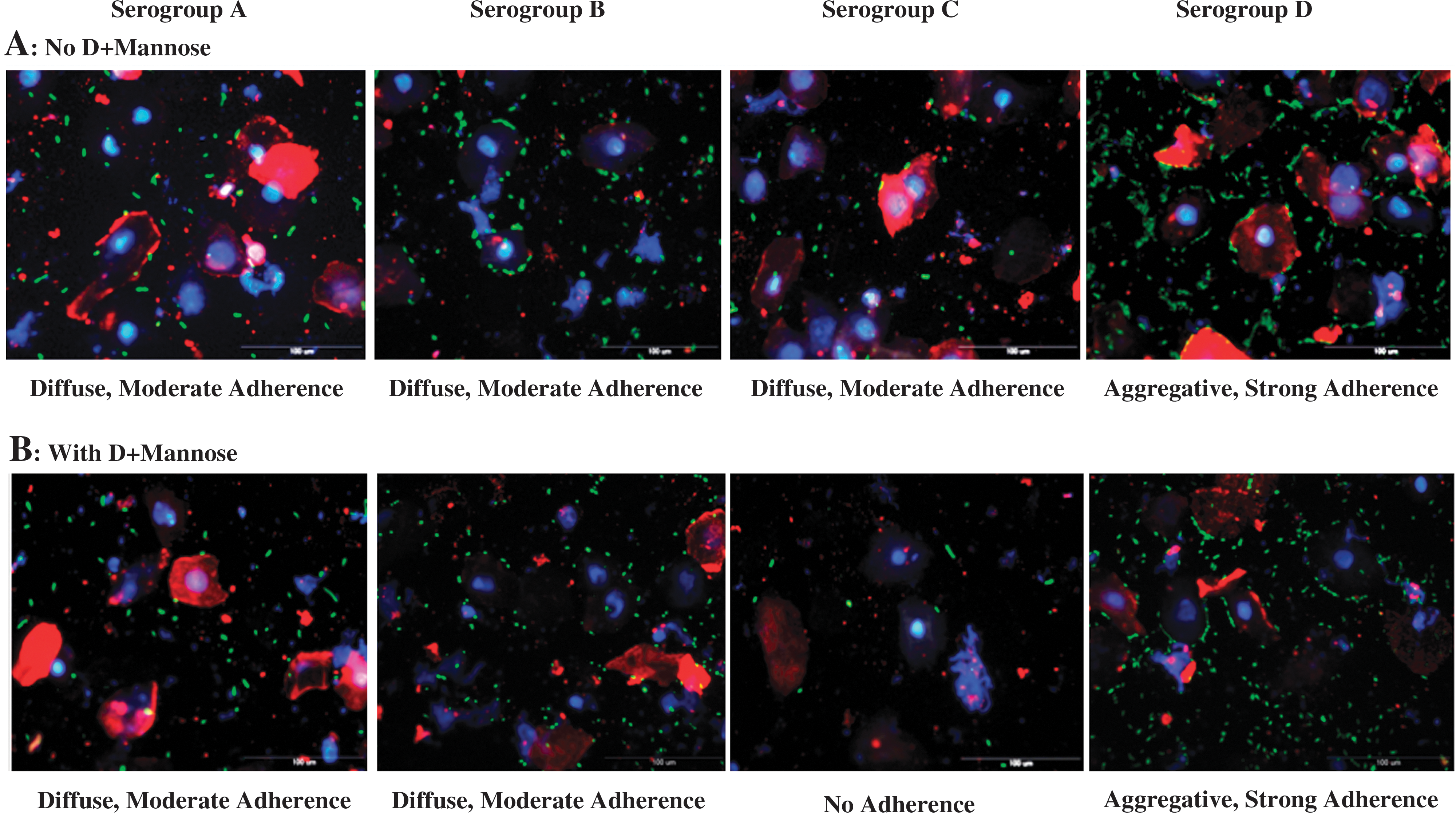
Adherence patterns of Shigella serogroups on recto-anal junction squamous epithelial (RSE) cells in the absence of D+mannose
The inclusion of D+mannose in the assay altered the adherence pattern of Shigella serogroup C in a significant manner. Serogroup C did not adhere to the RSE cells (non-adherent) in the presence of D+mannose (Fig. 1B and Table 1), and to highlight this change in the adherence pattern, data for serogroup C in the presence or absence of D+mannose is shown in Table 1. However, Shigella serogroups A, B, and D maintained their adherence patterns even in the presence of D+mannose (Fig. 1B and Table 1), with D+mannose dispersing only some of the bacterial aggregates in the background but not those on the RSE cells for serogroup D (Fig. 1B). Thus, by clearing non-specifically binding bacteria and altering the adherence of serogroup C, the adherence assay with D+mannose provided the most discerning adherence patterns for the Shigella serogroups tested that could be interpreted solely by qualitative analysis as well (Table 1).
Each trial had one slide per bacterial serogroup. Each slide in turn had eight separate RSE cells-bacteria mix spots on it; 10—20 well-dispersed RSE cells were evaluated per spot to get a total of 160 or 100.
Total number of RSE cells evaluated in each trial is shown in parentheses.
Number of bacteria adhering to each RSE cell is shown as a range of >10, 6–9, and 1–5. Number of RSE cells without bacteria is not shown.
MOI, multiplicity of infection.
The HEp-2 cell adherence assay, on the other hand, did not provide any distinguishing adherence patterns. All Shigella strains tested had similar diffuse adherence patterns in the presence (Fig. 2B) and absence of D+mannose (data not shown). No significant actin rearrangements were observed. This may have been due to the Shigella not invading these cells of human origin at the concentrations standardized for these assays, and instead adhering less intensely compared to the bovine RSE cells.
The bacteria/cell ratio was maintained at 10:1 for both the RSE and HEp-2 assays, as it provided optimal and consistent adherence results, similar to our previous observation (Kudva et al., 2011). The adherence patterns were consistent between repeat assays, as well as between different clinical strains of Shigella. No organisms were detected in the control slides without any added bacteria (data not shown). In the test slides that were treated with non-specific primary antibodies, no cross reactivity was observed with the anti-Salmonella antibodies (data not shown). E. coli K12 did not adhere to either epithelial cell types (Fig. 2A) (Kudva et al., 2011).
D+mannose was primarily included in both RSE and HEp-2 adherence assays to verify the role of Type 1 fimbriae in bacterial adherence to these cells (Ofek et al., 1978; Schaeffer et al., 1980; Rodrigues et al., 2009). These fimbriae appeared to play a role in the adherence of serogroups C to RSE cells only, as they showed altered adherence in the presence of D+mannose (Fig. 1). We are currently conducting experiments in cattle to see if Shigella adheres to RSE cells in vivo. If this is observed, we will dissect the mechanisms by which Shigella adhere to RSE cells. Our observations warrant both experimental and observational evaluation of Shigella serogroups A, B, and D carriage in cattle.
Footnotes
Acknowledgments
Technical assistance provided by Bryan Wheeler, Denise Chapman, and the animal caretakers at the NADC is acknowledged. We appreciate useful discussions with Dr. Michelle Crocheck, and the access to donor cattle for RAMS sampling and excess necropsy RAJ tissues from unrelated experiments. We especially acknowledge Dr. Mitchell Palmer, Dr. Judy Stabel, Dr. Manohar John, and Dr. Brian Brunelle for their insightful review of this manuscript.
Disclosure Statement
No competing financial interests exist.