
Abstract
Salmonella enterica serovar Infantis (Salmonella Infantis) is consistently isolated from broiler chickens, pigs, and humans worldwide. This study investigated 93 epidemiologically unrelated Salmonella Infantis strains isolated in Germany between 2005 and 2008 in respect to their transmission along the food chain. Various phenotypic and genotypic methods were applied, and the pathogenicity and resistance gene repertoire was determined. Phenotypically, 66% of the strains were susceptible to all 17 antimicrobials tested, while the others were almost all multidrug-resistant (two or more antimicrobial resistances), with different resistance profiles and preferentially isolated from broiler chickens. A number of phage types (PTs) were shared by strains from pigs, broiler chickens, and humans (predominated by PT 29). One, PT 1, was only detected in strains from pigs/pork and humans. Pulsed-field gel electrophoresis (PFGE) subdivided strains in seven different clusters, named A–G, consisting of 35 various XbaI profiles with coefficient of similarity values of 0.73–0.97. The majority of XbaI profiles were assigned to clusters A and C, and two predominant XbaI profiles were common in strains isolated from all sources investigated. Multi-locus sequence typing (MLST) analysis of selected strains representing the seven PFGE clusters revealed that they all belonged to ST32. The pathogenicity gene repertoire of 37 representative Salmonella Infantis strains analyzed by microarray was also identical. The resistance gene repertoire correlated perfectly with the phenotypic antimicrobial resistance profiles, and multidrug-resistant strains were associated with class 1 integrons. Overall, this study showed that two major closely related genotypes of Salmonella Infantis can transmit in Germany to humans through contaminated broiler meat or pork, and consequently presents a hazard for human health.
Introduction
Between the years 2000 and 2010, the German National Reference Laboratory for Salmonella (NRL-Salmonella) received on a routine basis an average of approximately 110 Salmonella Infantis strains (2.8% of all isolates) per year. Of these, 34% were isolated from broiler chickens or broiler meat and 11% from pig or pork. Other sources were cattle (7%), meat products of undefined origin, and, to a smaller degree, feeding stuff and environmental sources. Salmonella isolates from humans in the same time frame sent to the German National Reference Centre for Salmonellae and other Enterics were serotyped as Salmonella Infantis at an average of 84 cases per year (2.6% of all isolates) with increasing isolate numbers in 2009 (3.4%) and 2010 (4.2%). Increasing Salmonella Infantis isolation has been also recently reported in Israel (Gal-Mor et al., 2010) and Australia (Ross et al., 2008).
This study was conducted to gain a better understanding of the clonality of Salmonella Infantis and to understand the transmission of the serovar from broiler chickens and pigs to humans through food in Germany. For this purpose, 93 epidemiologically unrelated Salmonella Infantis strains isolated in Germany from different sources between 2005 and 2008 were investigated by phage typing, antimicrobial resistance, pulsed-field gel electrophoresis (PFGE), and partially multi-locus sequence typing (MLST). Additionally, the pathogenicity and antimicrobial resistance gene repertoire of the serovar was determined by microarray analysis.
Methods
Selection of strains
Altogether 93 Salmonella Infantis strains that had no obvious epidemiological link were used in this study (Table 1). Eight Salmonella Infantis strains were isolated from porcine lymph nodes during an EU Monitoring study in the years 2006 and 2007 on the prevalence of Salmonella in slaughter pigs (Anonymous, 2006). Six Salmonella Infantis strains were isolated from pork in the same time frame. From broiler chickens, 10 Salmonella Infantis strains were chosen from an EU Monitoring study in the years 2005 and 2006 on the prevalence of Salmonella in commercial broiler flocks of Gallus gallus (EFSA, 2007), and 20 strains were isolated from broiler meat in the same time frame. Only one strain was isolated in 2007 from plant-derived feeding stuff and was selected for this study. The food and feed strains selected were isolated nationwide from regional food diagnostic laboratories and sent to the National Reference Laboratory for Salmonella (NRL-BFR), Berlin, Germany for serotyping on a routine procedure. The remaining 48 Salmonella Infantis strains were isolated from epidemiologically unrelated human gastroenteritis cases between 2006 and 2008 selected from the strain collection of the National Reference Centre for Salmonellae and other Enterics (NRZ-RKI), Wernigerode, Germany. All strains were selected to represent various geographical origins in Germany as well as to cover different seasons. To our knowledge, none of the selected isolates were associated with foreign-acquired salmonellosis.
Strains for which pathogenicity gene repertoire was assessed by microarray.
Strains that were analyzed by multi-locus sequence typing (MLST).
PFGE, pulsed-field gel electrophoresis; n.t., non-typeable. For other abbreviations, see Methods section.
Serotyping
All strains were previously serotyped according to the White-Kauffmann-Le Minor scheme (Grimont and Weill, 2007) by slide agglutination with O- and H-antigen specific sera (Sifin Diagnostics, Berlin, Germany).
Phage typing
Phage typing was performed according to the phage typing scheme developed by Miller (2009). The phage typing system consisted of 17 typing phages defining 61 different phage types (PTs).
Antimicrobial susceptibility testing
Antimicrobial susceptibility of strains was tested against 17 antimicrobials or antimicrobial combinations by determining the minimum inhibitory concentration (MIC) using the Clinical and Laboratory Standards Institute (CLSI) broth micro-dilution method (CLSI, 2006) in combination with the semi-automatic Sensititre system (TREK Diagnostic Systems, Cleveland, OH). Breakpoints were applied as previously described (Schroeter et al., 2004). Antimicrobials tested were amoxicillin/clavulanic acid (AMC), ampicillin (AMP), chloramphenicol (CHL), ciprofloxacin (CIP), colistin (COL), florfenicol (FLO), gentamicin (GEN), kanamycin (KAN), neomycin (NEO), nalidixic acid (NAL), spectinomycin (SPE), streptomycin (STR), sulfamethoxazole (SMX), trimethoprim/sulfamethoxazole (SXT), tetracycline (TET), trimethoprim (TMP), and ceftiofur (XNL).
MLST
MLST was carried out as previously described (Kidgell et al., 2002), and the alleles and sequence types were assigned according to the MLST scheme at
PFGE
PFGE was carried out after digestion of genomic DNA with the restriction enzyme XbaI according to the Pulse-Net protocol (Ribot et al., 2006). Gel images were analyzed in BioNumerics v.5.1 (Applied Maths, Sint-Martens-Latem, Belgium) and compared by cluster analysis using Dice coefficient and Unweighted Pair Group Method with Averages (UPGMA; dendrogram type) with a position tolerance of 1.5% and optimization of 1.0%. Fragments that were smaller than 20 kb were not considered for cluster analysis.
DNA microarray typing
The DNA microarray typing used in this study was applied as previously described (Huehn et al., 2009a). Genomic DNA isolation was carried out using the DNeasy Blood and Tissue Kit (Qiagen GmbH, Hilden, Germany) according to the manufacturer's protocol with the addition of 25 μL Proteinase K instead of 20 μL and extended lysis for 3.5 h. The quality and quantity of DNA was measured spectrophotometrically, and a minimum of 4 μg of high-quality DNA was used for labeling.
Statistical analysis
To assess the discriminatory power of PFGE and phage typing Simpson's index of diversity (ID) and the 95% confidence intervals (CI) were calculated using the Comparing Partitions website (
Results
Antimicrobial resistance
Sixty-one of the 93 Salmonella Infantis strains (66%) were susceptible to all 17 antimicrobials tested, while two strains were mono-resistant to nalidixic acid or sulfamethoxazole (Table 1). One strain was resistant to STR and TET. Seven strains were multidrug-resistant to three antimicrobials (five different profiles) and 22 strains to four or more antimicrobials (nine different profiles). The percentage of resistant strains was significantly higher in those from broiler chickens or broiler meat origin (53%) compared to pig or pork (21%) or to human (25%; p<0.05). Seven out of 20 Salmonella Infantis strains isolated from broiler meat and one strain isolated from pork exhibited the antimicrobial resistance profile NAL STR TET SPE SMX, which was therefore the most prevalent profile.
Phage typing
Altogether 18 different PTs and one non-typeable strain were identified (ID 80.8 [95% CI, 73.9–87.6]). Thirty-seven of the Salmonella Infantis strains (40%) belonged to PT 29, followed by 14 of PT 1 (15%) and eight of PT 4 (9%) strains. Whereas PT 29 strains were mainly associated with those from broiler, broiler meat, and humans, PT 1 exclusively occurred in strains isolated from pig, pork, and humans. PT 4 was represented by strains from broiler, pig, and humans (Table 1).
Typing by PFGE
PFGE using XbaI restriction enzyme revealed 35 different XbaI profiles within the 93 strains analyzed (ID, 92.0 [95% CI, 88.6–95.4]) (Fig. 1). The main restriction profile was XINF10, found in 21 strains (23%) isolated from all sources except the feed strain. Four profiles (XINF09, XINF17, XINF21, and XINF25) were associated with human, pig, or broiler (Fig. 1). The remaining 30 profiles were associated with pig, pork, or human, often represented by one or two single strains. Seven different clusters (clusters A–G) were assigned after generating an UPGMA tree using the Dice similarity coefficient with coefficient of similarity (F) values of 0.73–0.97. Cluster A (F value of 0.85–0.97) comprised 53 of the 93 strains (57%) separated in 13 different XbaI profiles, including the most prominent restriction profile XINF10 (21 strains). Of these 21 XINF10 strains, 17 strains belonged to PT 29. The closely related profile XINF09 comprised eight additional strains assigned to PT 29. Cluster C (F value of 0.85–0.97) was represented by 31 strains (33%) comprising 13 profiles, with most predominant XINF17 profile (11 strains). Seven of the 11 XINF17 strains were assigned to PT 4. Other clusters were represented by only single strains (Fig. 1). The strain isolated from feed belonged to cluster D.

Unweighted Pair Group Method with Averages (UPGMA) dendrogram of pulsed-field gel electrophoresis (PFGE) profiles identified in 93 Salmonella Infantis strains after restriction with XbaI. Profiles were designated XINF01 to XINF35. The number of strains belonging to each source (total, pig, pork, broiler, broiler meat, and human) is shown on the right side. Assigned clusters A to G are indicated by brackets. *Strain with PFGE profile XINF29 was isolated from feed.
MLST
Eleven strains representing PFGE clusters A–G with XbaI types XINF02 (cluster A), XINF10 (A), XINF15 (B), XINF17 (C), XINF17, XINF21 and XINF28 (C), XINF29 (D), XINF31 (E), XINF33 (F), and XINF35 (G) were further typed by MLST, including seven housekeeping genes aroC, dnaN, hemD, hisD, purE, sucA, and thrA (Kidgell et al., 2002). All strains shared the same sequence type (ST) 32.
Microarray analysis of pathogenicity genes
Thirty-seven strains were selected for identification of 102 pathogenicity-associated genes by microarray analysis (Table 1). The selection represented diversity in resistance, phage, and PFGE types. All strains harbored the same pathogenicity array type. Genes targeted by the array and usually located within the Salmonella pathogenicity islands (SPIs) 1, 2, 4, and 5 were all present. In SPI-3, the genes rhuM and sugR encoding a putative cytoplasm protein and a putative ATP binding protein, respectively, were absent. Pathogenicity genes found in SPI-7 and the prophages Gifsy-1, Gifsy-2, Gifsy-3, Fels-1, and other prophages as well as genes located on the Salmonella virulence plasmid were completely absent. Additionally, signals for probes with specific sequences to genes cdtB (encoding cytolethal distending toxin), fhuA_Spa (outer membrane receptor, Salmonella Paratyphi A specific sequence), htrE (putative porin/fimbrial assembly protein), msgA (SsrB-regulator factor), sirA (regulator for invasion proteins), stkC (membrane usher protein), and trhH (pilus assembly protein) were absent.
In all strains, fimbrial markers agfA, bcfC, csgA, fimA, lpfD, safC, stbD, stdB, stfE, stiC, STM4595, and tcfA were present. Fimbrial markers pefA, prot6e, sefA/sefR, staA, stcC, steB, stgA, and stjB were absent.
Determination of resistance gene repertoire
Of the 37 strains investigated, 23 were susceptible, with none of the resistance genes analyzed present. Eleven out of the 14 multidrug-resistant strains tested were positive for int1 (DNA integrase 1), indicating an integron 1 cluster (Fig. 2). Ampicillin resistance was attributed to blaTEM1_like (encoding ß-lactamase) in all six strains tested. Trimethoprim resistance was associated with dfrA1-like (dihydrofolate reductase) except for one strain (08-03341) encoding dfrA14 confirmed by polymerase chain reaction (PCR) with primers previously described (Frech et al., 2003). This strain had also a sat1 (int2) gene (encoding a streptothricin acetyltransferase, usually linked to integrase 2). Streptomycin resistance was in eight of nine resistance profiles encoded by aadA1 (aminoglycoside-adenyltransferase). Tetracycline resistance was associated with tet(A), except in one profile (08-03346) where it was encoded by tet(B). Resistance to sulfmethoxazole was either encoded by sul1, sul2, or both genes.
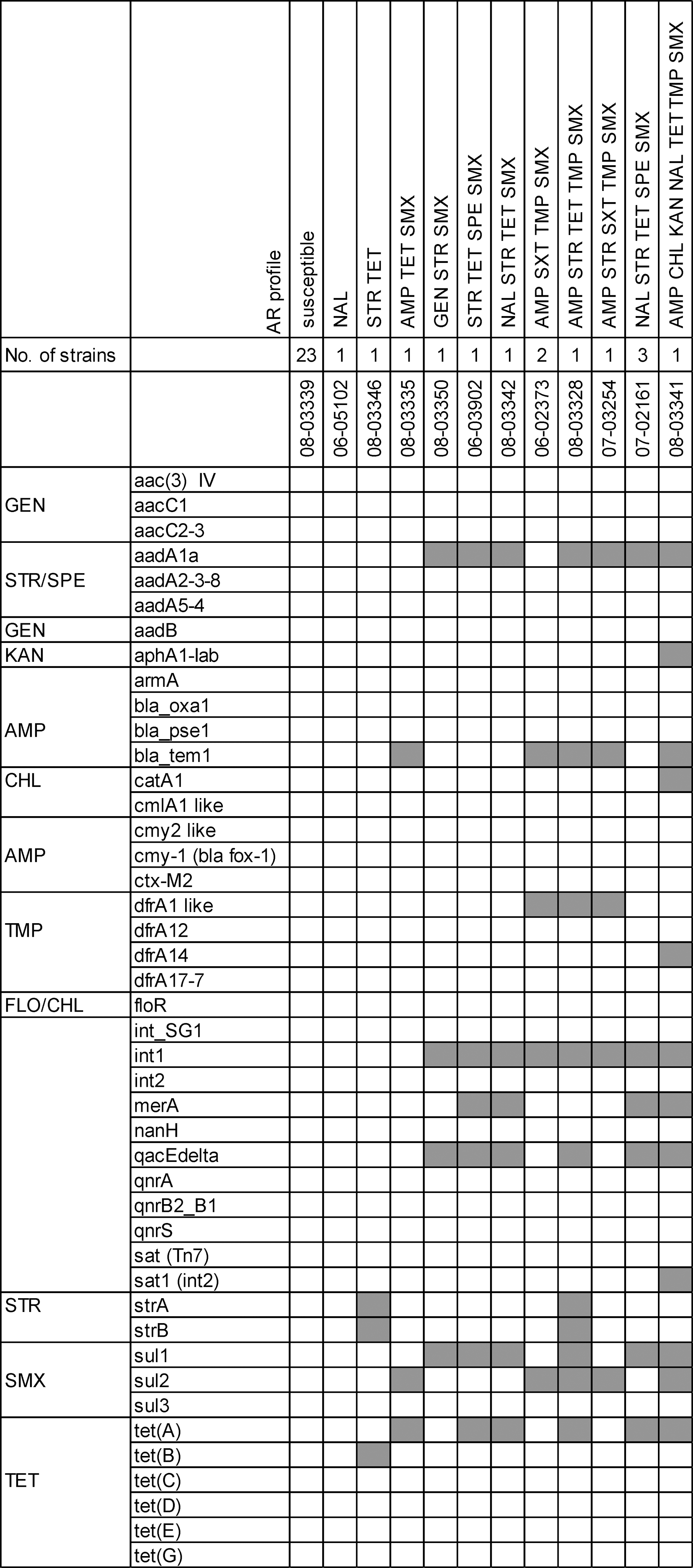
Resistance array types observed in 37 Salmonella Infantis strains tested. At the left, genes are indicated and grouped according to their particular antimicrobial resistance phenotype (for abbreviations, see Methods) or genetic resistance elements. At the top, specific antimicrobial resistance phenotypes analyzed, the number of strains harboring the particular antimicrobial resistance phenotype, and a corresponding Salmonella Infantis reference strain number are indicated. The hybridization result of each type is shown by row. A white box indicates the absence and a gray box the presence of the target sequence.
Characterization of clonal subgroups
Seven clusters were defined based on XbaI-PFGE typing results (Fig. 1). The two most prominent clusters comprised 53 strains (cluster A) and 31 strains (cluster C), respectively. Strains of cluster A belonged mainly to XINF10 (40%). Nine different PTs were found in cluster A. PT 29, with 34 strains (64%), was the predominant PT. Altogether 10 different PTs occurred in cluster C. PT 4 was, with seven strains, the most prominent. All PT 4 strains were correlated with XbaI profile XINF17.
Forty percent of the strains from cluster A and 29% from cluster C harbored antimicrobial resistance. The multidrug-resistant strains comprising profile NAL STR TET SPE SMX were all in cluster A located and correlated with XbaI profile XINF08 or XINF09 and PT 29.
The remaining PFGE clusters were represented by only a few or single strains isolated mainly from humans and harboring various properties in terms of antimicrobial resistance and PT.
Discussion
This study was conducted to determine the clonal structure among German Salmonella Infantis strains and to investigate the role of broiler meat and pork as vehicle in transmission of the serovar. The vast majority of strains tested were assigned to two XbaI-PFGE clusters A (61%) or C (30%). The predominant PFGE types within each of the two clusters were derived from strains isolated from primary production of pigs and broiler chickens, pork or broiler meat, and humans (XINF10, cluster A; XINF17, cluster C). Another PFGE type (XINF25) was identical between a pork and human strain, or between broiler and human strains (XINF21). The conclusion is that these Salmonella Infantis types spread in Germany from food-producing animals via the food chain to humans and cause infection. A number of other different XbaI-PFGE profiles were obtained with minor difference in band pattern often represented by single strains, especially those isolated from humans. It might be possible that these humans were infected sporadically by Salmonella Infantis from another source such as cattle or reptiles, which were not considered in this study. Alternatively, humans may have been infected abroad, but isolation data of the attending physicians did not give any hint of travel-related salmonellosis.
The study strongly indicates that Salmonella Infantis can be regarded as clonal serovar. One indication is that the PFGE band pattern variation among XbaI profiles in Salmonella Infantis strains is rather low. The lowest observed coefficient similarity (F) between strains was 0.72. Generally, a coefficient of similarity of more than 0.70 from PFGE types is indicative of a clonal relationship (Tenover et al., 1995; Goering, 2004). Other studies reported a similar high genetic relationship between Salmonella Infantis strains sampled over a longer period (Dunowska et al., 2007; Lindqvist and Pelkonen, 2007). Similar or identical PFGE patterns were also described in other studies from Japan (Noda et al., 2010), Hungary (Nogrady et al., 2007), Israel (Gal-Mor et al., 2010), Argentinia (Merino et al., 2005), and California (Gorski et al., 2011), indicating that the serovar is disseminated worldwide and possesses a highly clonal population structure. The clonality hypothesis is also supported by the observation that only one MLST type was identified in this study and all strains tested by DNA microarray harbored an identical pathogenicity gene array type. This array type was also previously observed in strains isolated from poultry or human in other European countries (Huehn et al., 2010). Consequently, strains analyzed in this study belong to the same clonal group, since the genetic variation is still too low to categorize them in different clonal lineages. Possibly, Salmonella Infantis has developed mechanisms protecting the serovar from major genetic rearrangements or horizontal genetic transfers. Another explanation is that the serovar has a recent ancestor and was yet not able to accumulate major evolutionary changes and their broad dissemination.
Of public health concern is the increasing isolation of multidrug-resistant Salmonella Infantis types in broiler chickens. In Hungary, an emerging multidrug-resistant subclone of Salmonella Infantis was identified frequently isolated from broiler chickens, broiler meat, and humans (Nogrady et al., 2007; Nogrady et al., 2008). This type exhibited characteristics that were also identified in strains from broiler meat and pork isolated in Germany. It was resistant to nalidixic acid, streptomycin (encoded by aadA1 in combination with a class 1 integron), sulphonamide (encoded by sul1), and tetracycline (encoded by tet(A)).The predominant pulsotype B2 reported from Hungarian isolates with this multidrug resistance was apparently identical with the one associated here (XINF08). Received Salmonella Infantis isolates from broiler meat based on a routine diagnostic at the NRL-Salmonella revealed that the phenotypic resistance type was recognized for the first time in 2000. Meanwhile, this multidrug-resistance type has been also established in German broiler chickens with a proportion of approximately 40% each year. It remains to be elucidated if other European countries were also affected. A recent study determining the antimicrobial resistance of Salmonella Infantis strains from Italy observed this type within a minority of strains (Dionisi et al., 2011). Similar multidrug-resistant types and their encoding resistance genes were described in broiler chicken from Japan (Asai et al., 2006; Shahada et al., 2006), but it is unclear how closely related this subclone is to the one described in this study. Another study determining 20 Turkish Salmonella Infantis strains isolated from broiler meat revealed a genotypic resistance pattern which was identical with one single strain (08-03341) isolated from humans in this study harboring aadA1, aph1, sul1, tet(A), dfrA5/dfrA14, and int1 (Abbasoglu and Akcelik, 2011). Several multidrug-resistant patterns comprised resistance against ampicillin (Table 1). In six different patterns tested, the ampicillin resistance was always encoded by bla TEM1-like (Fig. 2). Although ampicillin resistance was also reported from other countries, the responsible gene was often not analyzed. However, in Salmonella Infantis strains from Italy and Brazil, and in one strain from a Japanese study, ampicillin resistance was also encoded by bla TEM (Dionisi et al., 2011; Fonseca et al. 2006; Shahada et al., 2006). Possibly, this gene is responsible for ampicillin resistance in Salmonella Infantis. Another gene, bla CMY-2, which confers resistance to ampicillin and extended-spectrum cephalosporins, was described in Salmonella Infantis isolates from healthy pigs in Japan (Dahshan et al., 2010).
Conclusion
In conclusion, this study showed that, based on PFGE, two closely related genotypes of Salmonella Infantis were identified in Germany in isolates from pigs, broiler chickens, meat thereof, and humans. Consequently, those subgroups can be transmitted to humans via meat and are presumed to be a hazard for human health. One distinct multidrug-resistant subtype exhibiting resistance to NAL STR TET SPE and SMX is frequently isolated in Germany from broiler chickens and broiler meat, as well as from humans. To prevent this serovar and their multidrug-resistant variants from entering the food chain, control measurements should be enacted at the primary production location.
Footnotes
Acknowledgments
This work was funded by the Bundesministerium für Bildung und Forschung (BMBF), FBI-Zoo (01 KI 07123 [BFR] and 01 KI 07127 [RKI]). We thank Gabriele Berendonk, Manuela Jaber, Martha Brom, and Katharina Thomas for serotyping and antimicrobial resistance testing of strains.
Disclosure Statement
No competing financial interests exist.